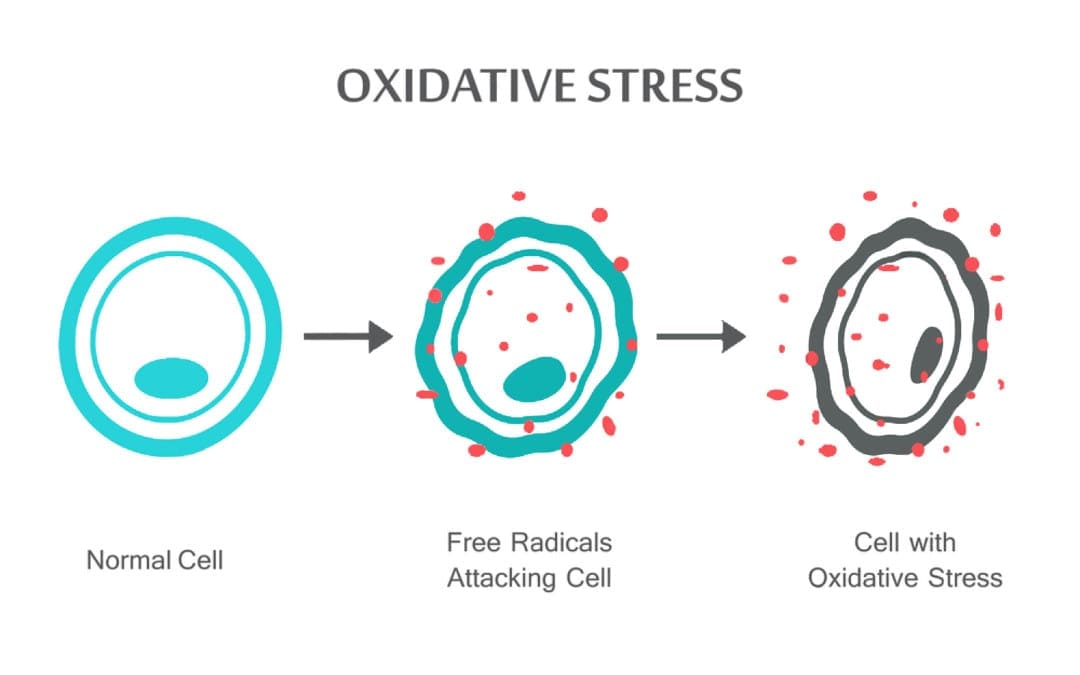
બેક ક્લિનિક ઓક્સિડેટીવ સ્ટ્રેસ ચિરોપ્રેક્ટિક અને કાર્યાત્મક દવા ટીમ. ઓક્સિડેટીવ તણાવને પ્રતિક્રિયાશીલ ઓક્સિજન (ફ્રી રેડિકલ) અને એન્ટીઑકિસડન્ટ સંરક્ષણના ઉત્પાદન વચ્ચેના સંતુલનમાં ખલેલ તરીકે વ્યાખ્યાયિત કરવામાં આવે છે. બીજા શબ્દોમાં કહીએ તો, તે મુક્ત રેડિકલના ઉત્પાદન અને એન્ટીઑકિસડન્ટો દ્વારા નિષ્ક્રિયકરણ દ્વારા હાનિકારક અસરોનો સામનો કરવા અથવા ડિટોક્સિફાય કરવાની શરીરની ક્ષમતા વચ્ચેનું અસંતુલન છે. ઓક્સિડેટીવ તણાવ શરીરમાં ઘણી પેથોફિઝીયોલોજીકલ પરિસ્થિતિઓ તરફ દોરી જાય છે. આમાં ન્યુરોડિજનરેટિવ રોગોનો સમાવેશ થાય છે, એટલે કે, પાર્કિન્સન રોગ, અલ્ઝાઈમર રોગ, જનીન પરિવર્તન, કેન્સર, ક્રોનિક ફેટીગ સિન્ડ્રોમ, નાજુક X સિન્ડ્રોમ, હૃદય અને રક્ત વાહિની વિકૃતિઓ, એથરોસ્ક્લેરોસિસ, હૃદયની નિષ્ફળતા, હૃદયરોગનો હુમલો અને બળતરા રોગો. ઓક્સિડેશન સંખ્યાબંધ સંજોગોમાં થાય છે:
કોષો ઊર્જા બનાવવા માટે ગ્લુકોઝનો ઉપયોગ કરે છે
રોગપ્રતિકારક તંત્ર બેક્ટેરિયા સામે લડે છે અને બળતરા પેદા કરે છે
શરીર પ્રદુષકો, જંતુનાશકો અને સિગારેટના ધુમાડાને ડિટોક્સિફાય કરે છે
આપણા શરીરમાં કોઈપણ સમયે લાખો પ્રક્રિયાઓ થઈ રહી છે જે ઓક્સિડેશનમાં પરિણમી શકે છે. અહીં કેટલાક લક્ષણો છે:
થાક
મેમરી નુકશાન અને અથવા મગજ ધુમ્મસ
સ્નાયુ અને અથવા સાંધાનો દુખાવો
ગ્રે વાળ સાથે કરચલીઓ
દૃષ્ટિમાં ઘટાડો
માથાનો દુખાવો અને અવાજ પ્રત્યે સંવેદનશીલતા
ચેપ માટે સંભાવિતતા
કાર્બનિક ખોરાકની પસંદગી અને તમારા વાતાવરણમાં ઝેરી પદાર્થોને ટાળવાથી મોટો ફરક પડે છે. આ, તણાવ ઘટાડવાની સાથે, ઓક્સિડેશન ઘટાડવામાં ફાયદાકારક બની શકે છે.
કોષ વિભાજન, બળતરા, રોગપ્રતિકારક કાર્ય, ઓટોફેજી અને તણાવ પ્રતિભાવ સહિત માનવ શરીરમાં આવશ્યક પ્રક્રિયાઓને નિયંત્રિત કરવા માટે સામાન્ય રીતે ઓક્સિડન્ટ્સ નિયંત્રિત રીતે ઉત્પન્ન થાય છે. જો કે, આ ઓક્સિડન્ટ્સનું અનિયંત્રિત ઉત્પાદન ફાળો આપી શકે છે ઓક્સિડેટીવ તણાવ, જે સેલ્યુલર કાર્યને અસર કરી શકે છે, જે ઝેરી, ક્રોનિક રોગ અને કેન્સરના વિકાસ તરફ દોરી જાય છે. માનવ શરીરની રક્ષણાત્મક એન્ટીઑકિસડન્ટ મિકેનિઝમ્સ મહત્વપૂર્ણ માર્ગોની શ્રેણી દ્વારા નિયંત્રિત થાય છે જે ઓક્સિડન્ટ્સ પ્રત્યે કોષની પ્રતિક્રિયાને નિયંત્રિત કરે છે. ન્યુક્લિયર ફેક્ટર એરિથ્રોઇડ 2-સંબંધિત પરિબળ, અન્યથા Nrf2 તરીકે ઓળખાય છે, તે ઓક્સિડન્ટ્સ માટે સેલ્યુલર પ્રતિકારનું ઉભરતું નિયમનકાર છે. નીચેના લેખનો હેતુ મિટોકોન્ડ્રીયલ કાર્યમાં Nrf2 ની ઉભરતી ભૂમિકાની ચર્ચા અને નિદર્શન કરવાનો છે.
અમૂર્ત
ટ્રાન્સક્રિપ્શન ફેક્ટર NF-E2 p45-સંબંધિત પરિબળ 2 (Nrf2; જનીન નામ NFE2L2) એન્ટીઑકિસડન્ટ, બળતરા વિરોધી, અને ડિટોક્સિફિકેશન વેલ એન્ઝાઇમ્સ સહિત સાયટોપ્રોટેક્ટીવ પ્રોટીનના વિવિધ નેટવર્ક્સના જનીન અભિવ્યક્તિને નિયંત્રિત કરીને તણાવની પરિસ્થિતિઓમાં અનુકૂલન અને અસ્તિત્વને મંજૂરી આપે છે. પ્રોટીન તરીકે કે જે ક્ષતિગ્રસ્ત મેક્રોમોલેક્યુલ્સના સમારકામ અથવા દૂર કરવામાં મદદ કરે છે. ગ્લુટાથિઓન, થિયોરેડોક્સિન અને એનએડીપીએચના જૈવસંશ્લેષણ, ઉપયોગ અને પુનર્જીવનનું નિયમન કરીને અને મિટોકોન્ડ્રિયા અને એનએડીપીએચ ઓક્સિડેઝ દ્વારા પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓના ઉત્પાદનને નિયંત્રિત કરીને સેલ્યુલર રેડોક્સ હોમિયોસ્ટેસિસની જાળવણીમાં Nrf2 નિર્ણાયક ભૂમિકા ધરાવે છે. હોમિયોસ્ટેટિક પરિસ્થિતિઓ હેઠળ, Nrf2 મિટોકોન્ડ્રીયલ મેમ્બ્રેન સંભવિત, ફેટી એસિડ ઓક્સિડેશન, શ્વસન માટે સબસ્ટ્રેટની ઉપલબ્ધતા (NADH અને FADH2/succinate) અને ATP સંશ્લેષણને અસર કરે છે. તણાવ અથવા વૃદ્ધિ પરિબળ ઉત્તેજનાની પરિસ્થિતિઓ હેઠળ, Nrf2 નું સક્રિયકરણ, અનકપ્લિંગ પ્રોટીન 3 ના ટ્રાન્સક્રિપ્શનલ અપગ્ર્યુલેશન દ્વારા મિટોકોન્ડ્રિયામાં વધેલા પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓના ઉત્પાદનનો સામનો કરે છે અને ન્યુક્લિયર રેસ્પિરેટરી ફેક્ટર 1 અને પેરોક્સિસોમ રિસેપ્ટેડ રિસેપ્ટર? કોએક્ટિવેટર 1?, તેમજ પ્યુરિન ન્યુક્લિયોટાઇડ બાયોસિન્થેસિસને પ્રોત્સાહન આપીને. ફાર્માકોલોજિકલ Nrf2 એક્ટિવેટર્સ, જેમ કે કુદરતી રીતે બનતું આઇસોથિયોસાયનેટ સલ્ફોરાફેન, મિટોકોન્ડ્રીયલ અભેદ્યતા સંક્રમણ છિદ્ર અને મિટોકોન્ડ્રીયલ સોજોના ઓક્સિડન્ટ-મધ્યસ્થી ઉદઘાટનને અટકાવે છે. જિજ્ઞાસાપૂર્વક, એક કૃત્રિમ 1,4-ડિફેનાઇલ-1,2,3-ટ્રાયઝોલ સંયોજન, જે મૂળરૂપે એનઆરએફ2 એક્ટિવેટર તરીકે રચાયેલ છે, તે મિટોફેજીને પ્રોત્સાહન આપવા માટે જોવા મળ્યું હતું, જેનાથી એકંદર મિટોકોન્ડ્રીયલ હોમિયોસ્ટેસિસમાં ફાળો આપે છે. આમ, Nrf2 એ મિટોકોન્ડ્રિયાની માળખાકીય અને કાર્યાત્મક અખંડિતતાને ટેકો આપવા માટે એક અગ્રણી ખેલાડી છે, અને આ ભૂમિકા ખાસ કરીને તણાવની પરિસ્થિતિઓમાં નિર્ણાયક છે.
સેલ્યુલર રેડોક્સ હોમિયોસ્ટેસિસ જાળવવામાં Nrf2 ની નિર્ણાયક ભૂમિકા છે.
Nrf2 મિટોકોન્ડ્રીયલ મેમ્બ્રેન સંભવિત અને ATP સંશ્લેષણને અસર કરે છે.
Nrf2 મિટોકોન્ડ્રીયલ ફેટી એસિડ ઓક્સિડેશનને પ્રભાવિત કરે છે.
Nrf2 મિટોકોન્ડ્રિયાની માળખાકીય અને કાર્યાત્મક અખંડિતતાને સમર્થન આપે છે.
જ્યારે માઇટોકોન્ડ્રીયલ કાર્ય સાથે ચેડા કરવામાં આવે ત્યારે Nrf2 એક્ટિવેટર્સ ફાયદાકારક અસરો ધરાવે છે.
પરિચય
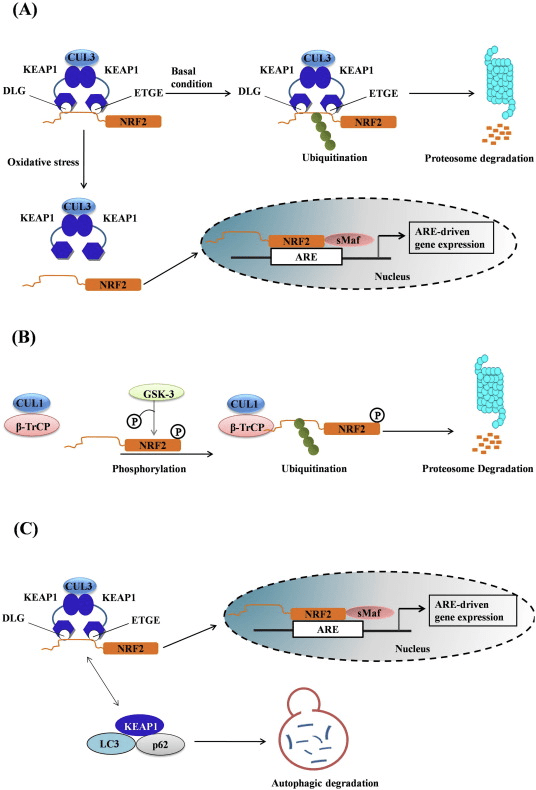
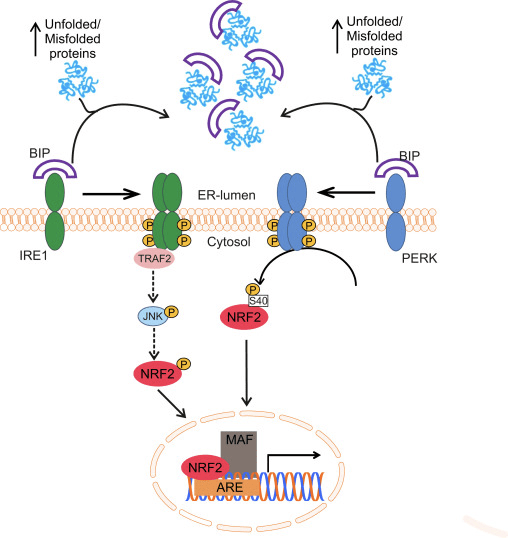
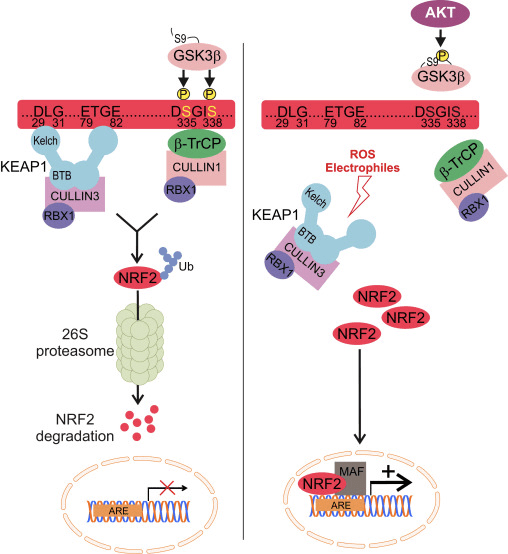
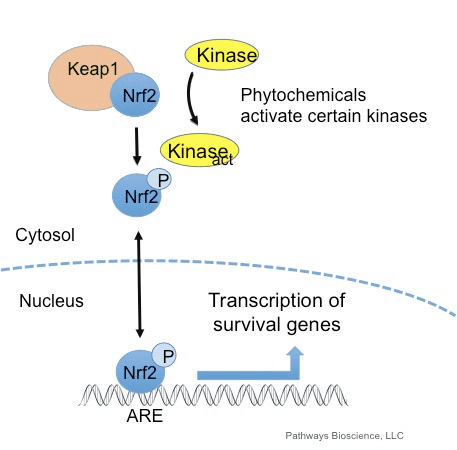
ટ્રાન્સક્રિપ્શન ફેક્ટર NF-E2 p45-સંબંધિત પરિબળ 2 (Nrf2; જનીન નામ NFE2L2) વિવિધ સાયટોપ્રોટેક્ટીવ પ્રવૃત્તિઓ સાથે જનીન એન્કોડિંગ પ્રોટીનના નેટવર્કની અભિવ્યક્તિને નિયંત્રિત કરે છે. Nrf2 પોતે મુખ્યત્વે પ્રોટીન સ્થિરતાના સ્તરે નિયંત્રિત થાય છે. મૂળભૂત પરિસ્થિતિઓમાં, Nrf2 એ અલ્પજીવી પ્રોટીન છે જે સતત સર્વવ્યાપકતા અને પ્રોટીસોમલ ડિગ્રેડેશનને આધિન છે. ત્યાં ત્રણ જાણીતી ubiquitin ligase સિસ્ટમો છે જે Nrf2 ના અધોગતિમાં ફાળો આપે છે. ઐતિહાસિક રીતે, શોધાયેલ Nrf2 નું પ્રથમ નકારાત્મક નિયમનકાર કેલ્ચ જેવું ECH-સંકળાયેલ પ્રોટીન 1 (Keap1) [1] હતું, જે Cullin 3 (Cul3)/Rbx1 ubiquitin ligase [2], [3], [4] માટે સબસ્ટ્રેટ એડેપ્ટર પ્રોટીન હતું. 1]. Keap2 સર્વવ્યાપકીકરણ અને પ્રોટીસોમલ ડિગ્રેડેશન માટે Nrf1 ને લક્ષ્ય બનાવવા માટે અત્યંત કાર્યક્ષમ ચક્રીય પદ્ધતિનો ઉપયોગ કરે છે, જે દરમિયાન Keap1 સતત પુનઃજનરેટ થાય છે, જે ચક્રને આગળ વધવા દે છે (ફિગ. 5A) [2]. Nrf3 પણ ગ્લાયકોજેન સિન્થેઝ કિનેઝ (GSK)1/?-TrCP-આધારિત Cul6-આધારિત ubiquitin ligase [7], [2] દ્વારા મધ્યસ્થી થતા અધોગતિને આધિન છે. તાજેતરમાં, એવું નોંધવામાં આવ્યું હતું કે, એન્ડોપ્લાઝમિક રેટિક્યુલમ તણાવની પરિસ્થિતિઓ દરમિયાન, E3 ubiquitin ligase Hrd1 [8] દ્વારા મધ્યસ્થી પ્રક્રિયામાં NrfXNUMX સર્વવ્યાપક અને અધોગતિ પામે છે.
આકૃતિ 1 Nrf1 ના Keap2-મધ્યસ્થી અધોગતિ માટે ચક્રીય અનુક્રમિક બંધન અને પુનર્જીવન મોડેલ. (A) Nrf2 ક્રમશઃ ફ્રી Keap1 ડાઇમર સાથે જોડાય છે: પ્રથમ તેના હાઇ-એફિનિટી ETGE (રેડ સ્ટીક્સ) બાઇન્ડિંગ ડોમેન દ્વારા અને પછી તેના લો-એફિનિટી DLG (બ્લેક સ્ટીક્સ) બાઇન્ડિંગ ડોમેન દ્વારા. પ્રોટીન સંકુલની આ રચનામાં, Nrf2 સર્વવ્યાપકતામાંથી પસાર થાય છે અને પ્રોટીસોમલ ડિગ્રેડેશન માટે લક્ષિત છે. ફ્રી Keap1 પુનઃજીવિત થાય છે અને નવા અનુવાદિત Nrf2 સાથે જોડાઈ શકે છે, અને ચક્ર ફરી શરૂ થાય છે. (B) ઈન્ડ્યુસર્સ (સફેદ હીરા) Keap1 (વાદળી લાકડીઓ) ના સેન્સર સિસ્ટીન સાથે પ્રતિક્રિયા આપે છે, જે રચનાત્મક પરિવર્તન તરફ દોરી જાય છે અને સબસ્ટ્રેટ એડેપ્ટર પ્રવૃત્તિને નબળી પાડે છે. ફ્રી Keap1 પુનઃજનરેટ થતું નથી, અને નવા સંશ્લેષિત Nrf2 એકઠા થાય છે અને ન્યુક્લિયસમાં સ્થાનાંતરિત થાય છે.
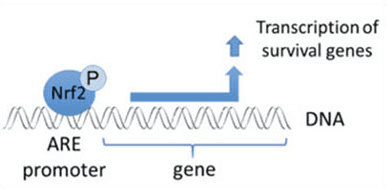
ubiquitin ligase સબસ્ટ્રેટ એડેપ્ટર પ્રોટીન તરીકે સેવા આપવા ઉપરાંત, Keap1 એ Nrf2 (ઇન્ડ્યુસર્સ તરીકે ઓળખાતા) [9] ના નાના-મોલેક્યુલ એક્ટિવેટર્સની વિશાળ શ્રેણી માટે સેન્સર પણ છે. ઇન્ડ્યુસર્સ Keap1 [2], [1] ની અંદર ચોક્કસ સિસ્ટીન અવશેષોને રાસાયણિક રીતે સંશોધિત કરીને અથવા Keap10:Nrf11 બંધનકર્તા ઇન્ટરફેસ [1], [2] ને સીધું વિક્ષેપિત કરીને Nrf12 ના Keap13-મધ્યસ્થી અધોગતિના ચક્રને અવરોધે છે. પરિણામે, Nrf2 અધોગતિ પામતું નથી, અને ટ્રાન્સક્રિપ્શન પરિબળ ન્યુક્લિયસ (ફિગ. 1B) માં સંચિત થાય છે અને સ્થાનાંતરિત થાય છે, જ્યાં તે નાના Maf પ્રોટીન સાથે હેટરોડીમર બનાવે છે; એન્ટીઑકિસડન્ટ-પ્રતિભાવ તત્વો સાથે જોડાય છે, તેના લક્ષ્ય જનીનોના અપસ્ટ્રીમ નિયમનકારી પ્રદેશો; અને ટ્રાન્સક્રિપ્શન શરૂ કરે છે [14], [15], [16]. Nrf2 લક્ષ્યોની બેટરીમાં ઝેનોબાયોટિક ચયાપચયના ઉત્સેચકો, એન્ટીઑકિસડન્ટ અને બળતરા વિરોધી કાર્યો સાથેના પ્રોટીન અને પ્રોટીસોમલ સબ્યુનિટ્સ તેમજ સેલ્યુલર રેડોક્સ હોમિયોસ્ટેસિસનું નિયમન કરતા પ્રોટીન અને મધ્યસ્થી મેટાબોલિઝમમાં ભાગ લેનારા પ્રોટીન સહિત વિવિધ સાયટોપ્રોટેક્ટીવ કાર્યો સાથે પ્રોટીનનો સમાવેશ થાય છે.
સેલ્યુલર રેડોક્સ હોમિયોસ્ટેસિસના મુખ્ય નિયમનકાર તરીકે Nrf2 નું કાર્ય વ્યાપકપણે ઓળખાય છે. ઘટેલા ગ્લુટાથિઓન (GSH) ના જૈવસંશ્લેષણમાં દર-મર્યાદિત પગલાને ઉત્પ્રેરક ઉત્પ્રેરક ઉત્પ્રેરક અને ?-ગ્લુટામિલ સિસ્ટીન લિગેસના નિયમનકારી સબ્યુનિટ્સ બંનેની જનીન અભિવ્યક્તિ, Nrf2 [17] દ્વારા સીધી રીતે નિયંત્રિત થાય છે. સિસ્ટમ xc-નું xCT સબ્યુનિટ, જે કોશિકાઓમાં સિસ્ટીન આયાત કરે છે, તે પણ Nrf2 [18] નું સીધું ટ્રાન્સક્રિપ્શનલ લક્ષ્ય છે. કોષમાં, સિસ્ટીન સિસ્ટીનમાં રૂપાંતરમાંથી પસાર થાય છે, જે GSH ના જૈવસંશ્લેષણ માટે પુરોગામી છે. GSH બાયોસિન્થેસિસમાં તેની ભૂમિકા ઉપરાંત, Nrf2 ગ્લુટાથિઓન રિડક્ટેઝ 1 [19], [20] ના સંકલિત ટ્રાન્સક્રિપ્શનલ નિયમન દ્વારા ગ્લુટાથિઓનને તેની ઘટેલી સ્થિતિમાં જાળવવા માટેનો ઉપાય પૂરો પાડે છે, જે NAD માંથી સમકક્ષ ઘટાડીને GSH માં ઓક્સિડાઇઝ્ડ ગ્લુટાથિઓન ઘટાડે છે. . જરૂરી NADPH ચાર મુખ્ય NADPH-જનરેટિંગ એન્ઝાઇમ્સ, મેલિક એન્ઝાઇમ 1 (ME1), આઇસોસીટ્રેટ ડિહાઇડ્રોજેનેઝ 1 (IDH1), ગ્લુકોઝ-6-ફોસ્ફેટ ડિહાઇડ્રોજેનેઝ (G6PD), અને 6-ફોસ્ફોગ્લુકોનેટ ડિહાઇડ્રોજેનેઝ (PGD) દ્વારા પ્રદાન કરવામાં આવે છે, જે તમામ છે. Nrf2 (ફિગ. 2) [21], [22], [23], [24] દ્વારા અંશતઃ ટ્રાન્સક્રિપ્શનલી નિયમન. જિજ્ઞાસાપૂર્વક, Nrf2 એલ્ડીહાઈડ ડીહાઈડ્રોજેનેઝ [25] ના સાયટોસોલિક, માઇક્રોસોમલ અને માઈટોકોન્ડ્રીયલ સ્વરૂપોના ઈન્ડ્યુસિબલ જનીન અભિવ્યક્તિને પણ નિયંત્રિત કરે છે, જે NAD(P)+ નો કોફેક્ટર તરીકે ઉપયોગ કરે છે, જે NAD(P)H ને જન્મ આપે છે. ખરેખર, એનએડીપીએચનું સ્તર અને એનએડીપીએચ/એનએડીપી+ રેશિયો એનઆરએફ2-નોકઆઉટ (એનઆરએફ2-કો) ઉંદરથી અલગ કરાયેલા ભ્રૂણ ફાઇબ્રોબ્લાસ્ટ્સમાં તેમના વાઇલ્ડ-ટાઇપ (ડબલ્યુટી) સમકક્ષોના કોષોની તુલનામાં નીચું છે, અને એનઆરએફ2 નોકડાઉન પર એનએડીપીએચ સ્તર ઘટે છે. રચનાત્મક રીતે સક્રિય Nrf2 [26] સાથે કેન્સર સેલ લાઇન. અપેક્ષા મુજબ, GSH નું સ્તર કોષોમાં ઓછું છે જેમાં Nrf2 વિક્ષેપિત થયો છે; તેનાથી વિપરીત, આનુવંશિક અથવા ફાર્માકોલોજિકલ માધ્યમ દ્વારા Nrf2 સક્રિયકરણ GSH અપગ્ર્યુલેશન તરફ દોરી જાય છે [27], [28], [29]. અગત્યની રીતે, Nrf2 થિયોરેડોક્સિન [30], [31], [32], થિયોરેડોક્સિન રીડક્ટેઝ 1 [28], [29], [32], [33] અને સલ્ફાયરેડોક્સિન [34] ના જનીન અભિવ્યક્તિને પણ નિયંત્રિત કરે છે, જે આવશ્યક છે. ઓક્સિડાઇઝ્ડ પ્રોટીન થિઓલ્સના ઘટાડા માટે.
આકૃતિ 2 ઝડપથી ફેલાતા કોષોના ચયાપચયમાં Nrf2 ની ભૂમિકા. Nrf2 એ બંને ઓક્સિડેટીવ આર્મ [એટલે કે, ગ્લુકોઝ-6-ફોસ્ફેટ ડિહાઈડ્રોજેનેઝ (G6PD) અને 6-ફોસ્ફોગ્લુકોનેટ ડિહાઈડ્રોજેનેઝ (PGD)] અને નોનઓક્સિડેટીવ આર્મ [એટલે કે, ટ્રાન્સલડોલેઝ 1 (TALDOLASE1) અને ટ્રાન્સકેટોલેસ 6 (TALDOLASE2) બંનેમાં જનીન એન્કોડિંગ એન્ઝાઇમ્સનું હકારાત્મક નિયમનકાર છે. TKT)] પેન્ટોઝ ફોસ્ફેટ માર્ગનો. G1PD અને PGD NADPH જનરેટ કરે છે. Nrf1 અન્ય બે NADPH-જનરેટિંગ એન્ઝાઇમ, મેલિક એન્ઝાઇમ 1 (ME1) અને આઇસોસીટ્રેટ ડિહાઇડ્રોજેનેઝ 2 (IDH2) ના જનીન અભિવ્યક્તિને પણ નિયંત્રિત કરે છે. ફોસ્ફોરીબોસિલ પાયરોફોસ્ફેટ એમીડોટ્રાન્સફેરેઝ (PPAT) ની જનીન અભિવ્યક્તિ, જે ડી નોવો પ્યુરીન બાયોસિન્થેટીક પાથવેમાં પ્રવેશને ઉત્પ્રેરિત કરે છે, તે પણ Nrf2 દ્વારા હકારાત્મક રીતે નિયમન કરવામાં આવે છે, જેમ કે મેથાઈલનેટેટ્રાહાઈડ્રોફોલેટ ડીહાઈડ્રોજેનેઝ 2 (MTHF6)ની ભૂમિકા સાથે એનઆરએફ 2 દ્વારા હકારાત્મક રીતે નિયમન કરવામાં આવે છે. ડી નોવો પ્યુરિન બાયોસિન્થેસિસ માટે એક-કાર્બન એકમો પ્રદાન કરે છે. Pyruvate kinase (PK) ને Nrf1 દ્વારા નકારાત્મક રીતે નિયમન કરવામાં આવે છે અને તે ગ્લાયકોલિટીક મધ્યવર્તી અને G6PD સાથે મળીને, પેન્ટોઝ ફોસ્ફેટ માર્ગ દ્વારા મેટાબોલાઇટ ચેનલિંગ અને ન્યુક્લીક એસિડ, એમિનો એસિડ અને ફોસ્ફોલિપિડ્સના સંશ્લેષણની તરફેણ કરે તેવી અપેક્ષા છે. Nrf6 એટીપી-સાઇટ્રેટ લાયઝ (સીએલ) ના જનીન અભિવ્યક્તિને નકારાત્મક રીતે નિયંત્રિત કરે છે, જે માઇટોકોન્ડ્રીયલ ઉપયોગ માટે સાઇટ્રેટની ઉપલબ્ધતા અથવા IDH6 માટે (આઇસોસીટ્રેટ દ્વારા) વધારી શકે છે. લાલ અને વાદળી અનુક્રમે હકારાત્મક અને નકારાત્મક નિયમન સૂચવે છે. મિટોકોન્ડ્રીયન ગ્રે રંગમાં બતાવવામાં આવે છે. મેટાબોલાઇટ સંક્ષેપ: G-6-P, ગ્લુકોઝ 1,6-ફોસ્ફેટ; F-1,6-P, ફ્રુક્ટોઝ 3-ફોસ્ફેટ; F-3-BP, ફ્રુક્ટોઝ 3-બિસ્ફોસ્ફેટ; GA-3-P, glyceraldehyde 6-phosphate; 6-પીજી, 6-ફોસ્ફોગ્લિસેરેટ; PEP, phosphoenolpyruvate; 6-પી-જીએલ, 5-ફોસ્ફોગ્લુકોનોલેક્ટોન; 5-પીજી, 5-ફોસ્ફોગ્લુકોનેટ; આર-1-પી, રિબ્યુલોઝ XNUMX-ફોસ્ફેટ; PRPP, XNUMX-ફોસ્ફોરીબોસિલ-?-XNUMX-પાયરોફોસ્ફેટ; THF, ટેટ્રાહાઇડ્રોફોલેટ; IMP, ઇનોસિન મોનોફોસ્ફેટ; એએમપી, એડેનોસિન મોનોફોસ્ફેટ; GMP, guanosine monophosphate.
સેલ્યુલર રેડોક્સ હોમિયોસ્ટેસિસના મુખ્ય નિયમનકાર તરીકે Nrf2 ની નિર્ણાયક ભૂમિકાને જોતાં, તે આશ્ચર્યજનક નથી કે, WT કોષોની તુલનામાં, પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓ (ROS) નું સ્તર કોષોમાં વધારે છે જેમાં Nrf2 વિક્ષેપિત થયો છે (Nrf2-KO) [35]. આ તફાવત ખાસ કરીને ઓક્સિડેટીવ તણાવ પેદા કરતા એજન્ટો સાથેના પડકાર પર પ્રહાર કરે છે. તદુપરાંત, Nrf2 માં ઉણપ ધરાવતા કોષો વિવિધ પ્રકારના ઓક્સિડન્ટ્સની ઝેરી અસર પ્રત્યે વધુ સંવેદનશીલ હોય છે અને Nrf2 ઇન્ડ્યુસર્સ દ્વારા તેને સુરક્ષિત કરી શકાતા નથી, જે સમાન પરિસ્થિતિઓ હેઠળ, WT કોષોને કાર્યક્ષમ અને લાંબા સમય સુધી રક્ષણ પૂરું પાડે છે [29], [36] , [37]. એકંદર સેલ્યુલર રેડોક્સ હોમિયોસ્ટેસિસ ઉપરાંત, Nrf2 એ મિટોકોન્ડ્રીયલ રેડોક્સ હોમિયોસ્ટેસિસની જાળવણી માટે પણ મહત્વપૂર્ણ છે. આમ, WT ની સરખામણીમાં, Keap1-KO માં કુલ મિટોકોન્ડ્રીયલ NADH પૂલ નોંધપાત્ર રીતે વધ્યો છે અને Nrf2-KO કોષોમાં નાટકીય રીતે ઘટાડો થયો છે [35].
લાઇવ સેલ ઇમેજિંગનો ઉપયોગ કરીને, અમે તાજેતરમાં WT, Nrf2-KO, અથવા Keap1-knockdown (Keap1-KD) ઉંદર [38] થી અલગ કરાયેલા પ્રાથમિક ગ્લિઓન્યુરોનલ કોકલ્ચર અને મગજની પેશીઓના ટુકડાઓમાં ROS ઉત્પાદનના દરોનું નિરીક્ષણ કર્યું છે. અપેક્ષા મુજબ, ROS ઉત્પાદનનો દર Nrf2-KO કોષો અને પેશીઓમાં તેમના WT સમકક્ષોની તુલનામાં ઝડપી હતો. જો કે, અમે અનપેક્ષિત અવલોકન કર્યું કે, WT ની તુલનામાં, Keap1-KD કોષોમાં પણ ROS ઉત્પાદનના ઊંચા દર છે, જો કે WT અને Keap1-KD જીનોટાઇપ્સ વચ્ચેના તફાવતની તીવ્રતા WT અને Nrf2-KO વચ્ચેના તફાવત કરતાં ઓછી હતી. . ત્યારબાદ અમે NOX2 અને NOX4 ના mRNA સ્તરોનું વિશ્લેષણ કર્યું, જે બે NADPH ઓક્સિડેઝ (NOX) isoforms ના ઉત્પ્રેરક સબ્યુનિટ્સ કે જે મગજની પેથોલોજીમાં સામેલ છે, અને જાણવા મળ્યું કે Nrf2 ની ઉણપની સ્થિતિમાં NOX2 નાટકીય રીતે વધે છે, જ્યારે NOX4 અપરેગ્યુલેટ થાય છે જ્યારે એનઆરએફ2 રચનાત્મક રીતે સક્રિય થાય છે, જોકે થોડી હદ સુધી. જથ્થાત્મક રીતે, મ્યુટન્ટ ઉંદરમાંથી કોષો અને પેશીઓમાં અપરેગ્યુલેશનની તીવ્રતા ROS ઉત્પાદન [38] માં અનુરૂપ વધારાને સમાંતર કરે છે. રસપ્રદ રીતે, Nrf2 માત્ર NADPH ઓક્સિડેઝનું નિયમન કરતું નથી, પરંતુ NADPH ઓક્સિડેઝ દ્વારા ઉત્પાદિત ROS Nrf2 ને સક્રિય કરી શકે છે, જેમ કે પલ્મોનરી એપિથેલિયલ કોષો અને કાર્ડિયોમાયોસાઇટ્સ [39], [40] માં દર્શાવવામાં આવ્યું છે. વધુમાં, ખૂબ જ તાજેતરના અભ્યાસે દર્શાવ્યું છે કે Nrf2 નું NADPH ઓક્સિડેઝ-આશ્રિત સક્રિયકરણ ક્રોનિક પ્રેશર ઓવરલોડ [41] દરમિયાન હૃદયમાં મિટોકોન્ડ્રીયલ નુકસાન અને કોષ મૃત્યુ સામે રક્ષણ માટે એક મહત્વપૂર્ણ અંતર્જાત મિકેનિઝમ બનાવે છે.
એનએડીપીએચ ઓક્સિડેઝની ઉત્પ્રેરક પ્રવૃત્તિ ઉપરાંત, મિટોકોન્ડ્રીયલ શ્વસન એ આરઓએસનો બીજો મુખ્ય અંતઃકોશિક સ્ત્રોત છે. મિટોકોન્ડ્રીયા-વિશિષ્ટ ચકાસણી MitoSOX નો ઉપયોગ કરીને, અમે પ્રાથમિક ગ્લિઓન્યુરોક્યુલ્યુલેટેડ ગ્લિઓન્યુરોકલમાં એકંદર આરઓએસ ઉત્પાદનમાં મિટોકોન્ડ્રીયલ મૂળના આરઓએસના યોગદાનની તપાસ કરી છે. WT, Nrf2-KO, અથવા Keap1-KD ઉંદર [38] માંથી. અપેક્ષા મુજબ, Nrf2-KO કોષોમાં WT કરતાં માઇટોકોન્ડ્રીયલ ROS ઉત્પાદનના ઊંચા દર હતા. એકંદર ROS ઉત્પાદન માટેના તારણો સાથે કરારમાં, Keap1-KD માં મિટોકોન્ડ્રીયલ ROS ઉત્પાદનના દર પણ WT કોષોની તુલનામાં વધુ હતા. મહત્વપૂર્ણ રીતે, રોટેનોન સાથે કોમ્પ્લેક્સ I ને અવરોધિત કરવાથી WT અને Keap1-KD બંને કોષોમાં મિટોકોન્ડ્રીયલ ROS ઉત્પાદનમાં નાટ્યાત્મક વધારો થયો હતો, પરંતુ Nrf2-KO કોષોમાં કોઈ અસર થઈ નથી. પાયરુવેટ ઉમેર્યા પછી WT કોષોમાં માઇટોકોન્ડ્રીયલ ROS ઉત્પાદનમાં અપેક્ષિત વધારાથી વિપરીત (NADH ની ઉપલબ્ધતા વધારવા માટે, માઇટોકોન્ડ્રીયલ મેમ્બ્રેન સંભવિતમાં વધારો કરવા અને શ્વસનને સામાન્ય બનાવવા), Nrf2-KO કોષોમાં ROS નું ઉત્પાદન ઘટ્યું. એકસાથે, આ તારણો ભારપૂર્વક સૂચવે છે કે, Nrf2 ની ગેરહાજરીમાં: (i) જટિલ I ની પ્રવૃત્તિ નબળી છે, (ii) જટિલ I ની ક્ષતિગ્રસ્ત પ્રવૃત્તિ સબસ્ટ્રેટની મર્યાદાને કારણે છે, અને (iii) જટિલની ક્ષતિગ્રસ્ત પ્રવૃત્તિ સંભવતઃ જટિલ II માંથી રિવર્સ ઇલેક્ટ્રોન પ્રવાહને કારણે, માઇટોકોન્ડ્રીયલ ROS ઉત્પાદનમાં વધારો થવાનું મુખ્ય કારણ I છે.
Nrf2 મિટોકોન્ડ્રીયલ મેમ્બ્રેન સંભવિત અને શ્વસનને અસર કરે છે
મિટોકોન્ડ્રીયલ મેમ્બ્રેન પોટેન્શિયલ (??m) એ મિટોકોન્ડ્રીયલ સ્વાસ્થ્ય અને કોષની મેટાબોલિક સ્થિતિનું સાર્વત્રિક સૂચક છે. તંદુરસ્ત કોષમાં, ??m એ મિટોકોન્ડ્રીયલ શ્વસન સાંકળ દ્વારા જાળવવામાં આવે છે. રસપ્રદ રીતે, એસ્ટ્રોજન રીસેપ્ટર-નેગેટિવ નોનટ્યુમોરિજેનિક માનવ સ્તન ઉપકલા MCF10A સેલ લાઇનમાં સંસ્કૃતિ-આધારિત પ્રોટીઓમિક્સ અભ્યાસમાં એમિનો એસિડ સાથે સ્થિર આઇસોટોપિક લેબલીંગ દર્શાવે છે કે માઇટોકોન્ડ્રીયલ ઇલેક્ટ્રોન ટ્રાન્સપોર્ટ ચેઇન ઘટક NDUFA4 અપરેગ્યુલેટેડ છે, જે એનડીયુએફએ (NDUFA2) દ્વારા અપરેગ્યુલેટેડ છે. જ્યારે Nrf2 નું આનુવંશિક અપરેગ્યુલેશન (Keap1 નોકડાઉન દ્વારા) સાયટોક્રોમ સી ઓક્સિડેઝ સબયુનિટ્સ COX2 અને COX4I1 [42] ના ડાઉનરેગ્યુલેશન તરફ દોરી જાય છે. દ્વિ-પરિમાણીય જેલ ઇલેક્ટ્રોફોરેસીસ અને મેટ્રિક્સ-આસિસ્ટેડ લેસર ડિસોર્પ્શન/આયનાઇઝેશન માસ સ્પેક્ટ્રોમેટ્રીનો ઉપયોગ કરીને લિવર પ્રોટીઓમના અભ્યાસમાં જાણવા મળ્યું છે કે Nrf2 એટીપી સિન્થેઝ સબ્યુનિટની અભિવ્યક્તિને નિયંત્રિત કરે છે? [43]. વધુમાં, મિટોકોન્ડ્રીયલ પ્રોટીન ડીજે-1, જે જટિલ I [44] ની પ્રવૃત્તિની જાળવણીમાં ભૂમિકા ભજવે છે, તે Nrf2 [45], [46] ને સ્થિર કરવા માટે નોંધવામાં આવ્યું છે, જોકે ફાર્માકોલોજીકલ અથવા આનુવંશિક સક્રિયકરણની ન્યુરોપ્રોટેક્ટીવ અસરો Nrf2 ના ડીજે-1 [47] થી સ્વતંત્ર છે. જો કે, મિટોકોન્ડ્રીયલ કાર્ય માટે આ અવલોકનોના પરિણામોની તપાસ કરવામાં આવી નથી.
Nrf2 ની ઉણપની પરિસ્થિતિઓ હેઠળ જટિલ I ની ક્ષતિગ્રસ્ત પ્રવૃત્તિ સાથેના કરારમાં, Nrf2-KO માઉસ એમ્બ્રીયોનિક ફાઇબ્રોબ્લાસ્ટ્સ (MEFs) અને સંસ્કારી પ્રાથમિક ગ્લિઓન્યુરોનલ કોશિકાઓમાં તેમના ડબલ્યુટી સમકક્ષો (ફિગ. 3, ઇન્સેટ) ની સરખામણીમાં બેઝલ ??m ઓછું છે. [35]. તેનાથી વિપરિત, જ્યારે Nrf2 આનુવંશિક રીતે બંધારણીય રીતે અપરેગ્યુલેટેડ હોય ત્યારે (Keap1 ના નોકડાઉન અથવા નોકઆઉટ દ્વારા) બેઝલ ??m વધારે હોય છે. જીનોટાઇપ્સમાં ??m માં આ તફાવતો સૂચવે છે કે Nrf2 ની પ્રવૃત્તિથી શ્વસન પ્રભાવિત થાય છે. ખરેખર, મૂળભૂત સ્થિતિમાં ઓક્સિજન વપરાશના મૂલ્યાંકનથી જાણવા મળ્યું છે કે, WT ની સરખામણીમાં, Nrf2-KO અને Keap1-KO MEF માં ઓક્સિજનનો વપરાશ અનુક્રમે ~50 અને ~35% ઓછો છે.
આકૃતિ 3 Nrf2 ની ઉણપની શરતો હેઠળ સમાધાન કરાયેલ મિટોકોન્ડ્રીયલ કાર્ય માટે સૂચિત પદ્ધતિ. (1) ME1, IDH1, G6PD, અને PGD ના ઘટેલા સ્તરના પરિણામે NADPH સ્તર નીચાં આવે છે. (2) GSH નું સ્તર પણ ઓછું છે. (3) ME1 ની ઓછી પ્રવૃત્તિ મિટોકોન્ડ્રિયામાં પ્રવેશતા પાયરુવેટના પૂલને ઘટાડી શકે છે. (4) NADH ની ઉત્પત્તિ ધીમી છે, જે જટિલ I ની નબળી પ્રવૃત્તિ તરફ દોરી જાય છે અને મિટોકોન્ડ્રીયલ ROS ઉત્પાદનમાં વધારો કરે છે. (5) માઇટોકોન્ડ્રીયલ પ્રોટીનમાં FAD થી FADH2 નો ઘટાડો પણ ઘટે છે, FADH2 થી UbQ અને જટિલ III માં ઇલેક્ટ્રોન પ્રવાહને ઘટાડે છે. (6) UbQH2 ની ધીમી રચના સુસીનેટ ડીહાઈડ્રોજેનેઝની એન્ઝાઇમ પ્રવૃત્તિને ઘટાડી શકે છે. (7) ROS ના વધેલા સ્તરો જટિલ II ની પ્રવૃત્તિને વધુ અવરોધે છે. (8) ફેટી એસિડ ઓક્સિડેશનની ઓછી કાર્યક્ષમતા મિટોકોન્ડ્રીયલ શ્વસન માટે સબસ્ટ્રેટની ઉપલબ્ધતામાં ઘટાડો કરે છે. (9) ઓક્સિડેટીવ ફોસ્ફોરીલેશનમાં ઘટતા ATP ઉત્પાદન માટે વળતરની પદ્ધતિ તરીકે ગ્લાયકોલીસીસને ઉન્નત કરવામાં આવે છે. (10) એટીપી સિન્થેઝ જાળવવા માટે વિપરીત રીતે કાર્ય કરે છે. લાલ અને વાદળી અનુક્રમે અપરેગ્યુલેશન અને ડાઉનરેગ્યુલેશન સૂચવે છે. બોક્સ પ્રાયોગિક પુરાવાઓની ઉપલબ્ધતા દર્શાવે છે. ઇનસેટ WT અને Nrf2-KO કોર્ટિકલ એસ્ટ્રોસાઇટ્સના મિટોકોન્ડ્રિયાની છબીઓ દર્શાવે છે જે પોટેન્ટિઓમેટ્રિક ફ્લોરોસન્ટ પ્રોબ ટેટ્રામેથિલરોડામાઇન મિથાઈલ એસ્ટર (TMRM; 25 nM) દ્વારા વિઝ્યુઅલાઈઝ કરવામાં આવી છે. સ્કેલ બાર, 20 �m.
જીનોટાઇપ્સમાં ??m અને શ્વસનમાં આ તફાવતો મિટોકોન્ડ્રીયલ શ્વસન માટે સબસ્ટ્રેટના ઉપયોગના દર દ્વારા પ્રતિબિંબિત થાય છે. ટ્રાઇકાર્બોક્સિલિક એસિડ (TCA) ચક્ર (મેલેટ/પાયરુવેટ, જે બદલામાં કોમ્પ્લેક્સ I સબસ્ટ્રેટ NADH) અથવા મિથાઈલ સસીનેટ, કોમ્પ્લેક્સ II માટે સબસ્ટ્રેટના ઉત્પાદનમાં વધારો કરે છે, માટે સબસ્ટ્રેટનો ઉપયોગ બંને WTમાં ??m માં એક પગથિયાંની વૃદ્ધિનું કારણ બને છે. અને Keap1-KD ચેતાકોષો, પરંતુ Keap1-KD કોષોમાં વૃદ્ધિનો દર વધારે છે. વધુ મહત્ત્વની વાત એ છે કે, આ TCA ચક્ર સબસ્ટ્રેટના પ્રતિભાવના આકાર બે જીનોટાઇપ્સ વચ્ચે અલગ છે, જેમાં સબસ્ટ્રેટ ઉમેર્યા પછી Keap1-KD કોષોમાં ??m માં ઝડપી વધારો, ઉચ્ચપ્રદેશને બદલે ઝડપી ડ્રોપ દ્વારા અનુસરવામાં આવે છે, જે અસામાન્ય રીતે સૂચવે છે. ઝડપી સબસ્ટ્રેટ વપરાશ. આ તારણો WT MEF ની સરખામણીમાં Keap50-KO માં [U-70C1] ગ્લુકોઝના 13-h પલ્સ પછી જોવા મળેલા મલેટ, પાયરુવેટ અને સક્સીનેટના ઘણા નીચા (6�1% દ્વારા) સ્તરો સાથે નજીકના કરારમાં છે. કોષો [24]. Nrf2-KO ચેતાકોષોમાં, માત્ર પાયરુવેટ ??m વધારવામાં સક્ષમ છે, જ્યારે મેલેટ અને મિથાઈલ સસીનેટ હળવા વિધ્રુવીકરણનું કારણ બને છે. માઇટોકોન્ડ્રીયલ સબસ્ટ્રેટ ઉત્પાદન પર Nrf2 ની અસર એ મુખ્ય પદ્ધતિ હોય તેવું લાગે છે જેના દ્વારા Nrf2 મિટોકોન્ડ્રીયલ કાર્યને અસર કરે છે. માઇટોકોન્ડ્રીયલ NADH રેડોક્સ ઇન્ડેક્સ (કોમ્પ્લેક્સ I દ્વારા NADH ના વપરાશ અને TCA ચક્રમાં NADPH ના ઉત્પાદન વચ્ચેનું સંતુલન) તેમના WT સમકક્ષોની સરખામણીમાં Nrf2-KO કોષોમાં નોંધપાત્ર રીતે નીચું છે, અને વધુમાં, પુલના પુનર્જીવનના દરો. જટિલ IV ના નિષેધ પછી NADH અને FADH2 (NACN ના ઉપયોગ દ્વારા) મ્યુટન્ટ કોશિકાઓમાં ધીમી છે.
મ્યુરિન મગજ અને યકૃતથી અલગ પડેલા મિટોકોન્ડ્રિયામાં, જટિલ I અથવા જટિલ II માટે સબસ્ટ્રેટની પૂરકતા જ્યારે Nrf2 સક્રિય થાય છે ત્યારે ઓક્સિજન વપરાશના દરને વધુ મજબૂત રીતે વધે છે અને જ્યારે Nrf2 વિક્ષેપિત થાય છે ત્યારે ઓછી કાર્યક્ષમતાથી [35]. આમ, ડબલ્યુટીની તુલનામાં કેપ1-કેડીમાં મેલેટ ઓક્સિજન વપરાશના ઊંચા દરને પ્રેરિત કરે છે, પરંતુ તેની અસર Nrf2-KO મિટોકોન્ડ્રિયામાં નબળી છે. તેવી જ રીતે, રોટેનોનની હાજરીમાં (જ્યારે જટિલ I અટકાવવામાં આવે છે), સસીનેટ WT ની તુલનામાં Keap1-KD માં વધુ પ્રમાણમાં ઓક્સિજન વપરાશને સક્રિય કરે છે, જ્યારે Nrf2-KO મિટોકોન્ડ્રિયામાં પ્રતિભાવ ઓછો થાય છે. વધુમાં, Nrf2-KO પ્રાથમિક ચેતાકોષીય સંસ્કૃતિઓ અને ઉંદર જટિલ II અવરોધકો 3-નાઇટ્રોપ્રોપિયોનિક એસિડ અને મેલોનેટની ઝેરીતા પ્રત્યે વધુ સંવેદનશીલ છે, જ્યારે Nrf2-ઓવરએક્સપ્રેસિંગ એસ્ટ્રોસાઇટ્સનું ઇન્ટ્રાસ્ટ્રિયાટલ ટ્રાન્સપ્લાન્ટેશન રક્ષણાત્મક છે [48], [49]. એ જ રીતે, Nrf2-KO ઉંદરો વધુ સંવેદનશીલ હોય છે, જ્યારે Nrf2 નું આનુવંશિક અથવા ફાર્માકોલોજિકલ સક્રિયકરણ 1-મિથાઈલ-4-ફિનાઈલ-1 માં જટિલ I અવરોધક 4-મિથાઈલ-1,2,3,6-ફેનિલપાયરિડિનિયમ આયનને કારણે ન્યુરોટોક્સિસિટી સામે રક્ષણાત્મક અસરો ધરાવે છે, પાર્કિન્સન્સ રોગનું 49-ટેટ્રાહાઇડ્રોપ્રાયરીડિન પ્રાણી મોડેલ [50], [51], [52], [53], [54], [55], [56], [57], [58], [59], [60], [61], [XNUMX].
શ્વસન નિયંત્રણ ગુણોત્તર (RCR), રાજ્ય 3 (ADP-ઉત્તેજિત) અને રાજ્ય 4 શ્વસનનો ગુણોત્તર (કોઈ ADP હાજર નથી), Nrf2 ની ગેરહાજરીમાં ઘટાડો થયો છે, પરંતુ RCR Keap1-KD અને WT મિટોકોન્ડ્રિયા [35 વચ્ચે સમાન છે. ]. જેમ કે આરસીઆર એ ઓક્સિડેટીવ ફોસ્ફોરાયલેશન માટે મિટોકોન્ડ્રીયલ શ્વસન સાંકળ પ્રવૃત્તિના જોડાણની ડિગ્રીનો સંકેત છે, આ શોધ સૂચવે છે કે કેપ1-કેડી મિટોકોન્ડ્રિયામાં શ્વસનનો ઉચ્ચ દર ઓક્સિડેટીવ ફોસ્ફોરીલેશનના જોડાણને કારણે નથી. તે વધુમાં સૂચવે છે કે જ્યારે Nrf2 સક્રિય થાય છે ત્યારે ઓક્સિડેટીવ ફોસ્ફોરાયલેશન વધુ કાર્યક્ષમ હોય છે. Keap1-KD મિટોકોન્ડ્રિયામાં શ્વસનનો ઊંચો દર મિટોકોન્ડ્રીયલ આરઓએસ ઉત્પાદનના ઊંચા સ્તરો [૩૮] સાથે સુસંગત છે કારણ કે ઊંચા શ્વસન દર ઈલેક્ટ્રોન લીકમાં વધારો કરી શકે છે. જો કે, ઓક્સિડેટીવ તણાવની પરિસ્થિતિઓમાં, વધેલા ROS ઉત્પાદનનો Nrf38-આશ્રિત ટ્રાન્સક્રિપ્શનલ અપગ્ર્યુલેશન ઓફ અનકપ્લીંગ પ્રોટીન 2 (UCP3) દ્વારા પ્રતિકાર કરવામાં આવે છે, જે મિટોકોન્ડ્રીયલ આંતરિક પટલના પ્રોટોન વહનને વધારે છે અને પરિણામે સુપરઓક્સાઇડનું ઉત્પાદન ઘટે છે [3]. ખૂબ જ તાજેતરમાં, એવું દર્શાવવામાં આવ્યું હતું કે લિપિડ પેરોક્સિડેશન પ્રોડક્ટ 62-હાઇડ્રોક્સી-4-નોનેનલ કાર્ડિયોમાયોસાઇટ્સમાં UCP2 ના Nrf2-આશ્રિત અપગ્ર્યુલેશનમાં મધ્યસ્થી કરે છે; ઓક્સિડેટીવ સ્ટ્રેસ જેવી કે ઇસ્કેમિયા રીપરફ્યુઝન [3] દરમિયાન રક્ષણ માટે આ ખાસ કરીને મહત્વનું હોઈ શકે છે.
Nrf2 ઓક્સિડેટીવ ફોસ્ફોરીલેશનની કાર્યક્ષમતા અને ATP ના સંશ્લેષણને અસર કરે છે
શ્વસન પર Nrf2 ની અસર સાથેના કરારમાં, મગજ અને યકૃતના મિટોકોન્ડ્રિયામાં, Nrf2 ની ઉણપ ઓક્સિડેટીવ ફોસ્ફોરાયલેશનની કાર્યક્ષમતામાં ઘટાડો કરે છે (એડીપી અને ઓક્સિજનના ગુણોત્તર દ્વારા અનુમાનિત કરવામાં આવે છે, જે ATP સંશ્લેષણ માટે વપરાય છે), જ્યારે Nrf2 સક્રિયકરણ (Keap1) -KD) ની વિપરીત અસર છે [35]. WT ની સરખામણીમાં, Nrf2 ના રચનાત્મક અપગ્ર્યુલેશન સાથેના કોષોમાં ATP સ્તર નોંધપાત્ર રીતે વધારે છે અને જ્યારે Nrf2 નીચે પછાડવામાં આવે છે [64] અથવા વિક્ષેપિત થાય છે [35]. વધુમાં, ઓક્સિડેટીવ ફોસ્ફોરીલેશન (ઓલિગોમાસીન) અથવા ગ્લાયકોલીસીસ (આયોડોએસેટિક એસિડ) ના અવરોધકોના ઉપયોગથી જાણવા મળ્યું છે કે Nrf2 કોષો ATP ઉત્પન્ન કરવાની રીતને બદલે છે. આમ, WT ચેતાકોષોમાં, oligomycin ATP માં સંપૂર્ણ ઘટાડાનું કારણ બને છે અને iodoacetic એસિડની વધુ અસર થતી નથી. નોંધપાત્ર રીતે, Nrf2-KO કોષોમાં, oligomycin ATP સ્તરમાં વધારો કરે છે, જે પછી ધીમે ધીમે, પરંતુ સંપૂર્ણપણે, iodoacetic એસિડ દ્વારા ક્ષીણ થઈ જાય છે, જે દર્શાવે છે કે Nrf2 ની ગેરહાજરીમાં, ગ્લાયકોલિસિસ, અને ઓક્સિડેટીવ ફોસ્ફોરીલેશન નહીં, એટીપી ઉત્પાદનનો મુખ્ય સ્ત્રોત છે. રસપ્રદ વાત એ છે કે, Keap1-KD કોષોમાં ઓક્સિડેટીવ ફોસ્ફોરાયલેશનની કાર્યક્ષમતા વધી હોવા છતાં, ઓલિગોમાસીન ઉમેરવાથી ATP સ્તરોમાં ~80% ઘટાડો થાય છે, અને આયોડોએસેટિક એસિડ વધુ ~20% ઘટાડાનું કારણ બને છે. આમ, Nrf2 ની ઉણપ અથવા તેની રચનાત્મક સક્રિયકરણ ઓક્સિડેટીવ ફોસ્ફોરીલેશનના યોગદાનને ઘટાડે છે અને ATP ના સંશ્લેષણ તરફ ગ્લાયકોલીસીસના યોગદાનને વધારે છે. આ અસર ખાસ કરીને ઉચ્ચારવામાં આવે છે જ્યારે Nrf2 ગેરહાજર હોય અને માધ્યમ [35] માં ગ્લુકોઝની હાજરી અને ગ્લાયકોલિટીક મધ્યવર્તી (G-6-P, F-6-P) ના વધેલા સ્તરો પર ??m ની અવલંબન સાથે સુસંગત હોય. , ડાયહાઇડ્રોક્સાયસેટોન ફોસ્ફેટ, પાયરુવેટ અને લેક્ટેટ) Nrf2 [24] ના નોકડાઉન પછી.
ઓલિગોમાસીન દ્વારા F1F0-ATPase ના નિષેધ પછી ATP સ્તરમાં વધારો સૂચવે છે કે Nrf2 ની ગેરહાજરીમાં, F1F0-ATPase ATPase તરીકે કાર્ય કરે છે અને ATP સિન્થેઝ તરીકે નહીં, એટલે કે, તે વિપરીત રીતે કાર્ય કરે છે. પ્રવૃતિમાં આ પ્રકારનું વિપરિત સંભવતઃ આંતરિક માઇટોકોન્ડ્રીયલ પટલમાં પ્રોટોનને પમ્પ કરવાની જરૂરિયાતને પ્રતિબિંબિત કરે છે ??m, જે આ અંગની કાર્યાત્મક અખંડિતતા માટે નિર્ણાયક છે. F1F0-ATPase ના કાર્યની ઉલટાવી પણ Nrf2-KO કોશિકાઓ પર ઓલિગોમાસીન વહીવટ પર અવલોકન કરાયેલ મિટોકોન્ડ્રીયલ વિધ્રુવીકરણ દ્વારા પુરાવા મળે છે, જે તેમના WT અથવા Keap1-ઉણપ ધરાવતા સમકક્ષો [35] માં થતા હાયપરપોલરાઇઝેશનથી તદ્દન વિપરીત છે. એકંદરે, એવું લાગે છે કે Nrf2 ની ઉણપની સ્થિતિમાં ATP મુખ્યત્વે ગ્લાયકોલીસીસમાં ઉત્પન્ન થાય છે, અને આ ATP પછી F1F0-ATPase દ્વારા ??m જાળવવા માટે આંશિક રીતે ઉપયોગમાં લેવાય છે.
Nrf2 મિટોકોન્ડ્રીયલ ફેટી એસિડ ઓક્સિડેશનને વધારે છે
??m પર Nrf2 ની ઉણપની અસર ખાસ કરીને ઉચ્ચારવામાં આવે છે જ્યારે કોષો ગ્લુકોઝ વિના માધ્યમમાં ઉકાળવામાં આવે છે, અને WT કોષો [50] ની સરખામણીમાં Nrf2-KO માં ??m ~35% ઓછું છે. ગ્લુકોઝની વંચિતતાની સ્થિતિમાં, મિટોકોન્ડ્રીયલ ફેટી એસિડ ઓક્સિડેશન (FAO) એ શ્વસન અને ઓક્સિડેટીવ ફોસ્ફોરાયલેશન માટે સબસ્ટ્રેટનું મુખ્ય પ્રદાતા છે, જે સૂચવે છે કે Nrf2 FAO ને અસર કરી શકે છે. ખરેખર, FAO ની કાર્યક્ષમતા બંને લોંગ-ચેઈન (C16:0) સંતૃપ્ત ફેટી એસિડ પામમેટિક એસિડ અને શોર્ટ-ચેન (C6:0) હેક્સાનોઈક એસિડ માટે Keap1-KO MEFs અને અલગ હૃદય અને યકૃત મિટોકોન્ડ્રિયામાં તેમના કરતાં વધુ છે. WT સમકક્ષ, જ્યારે તે Nrf2-KO કોષો અને મિટોકોન્ડ્રિયા [65] માં ઓછું છે. આ અસરો મનુષ્યો માટે પણ અત્યંત સુસંગત છે: ખરેખર, TCA ચક્રની પ્રવૃત્તિ સાથે FAO ના વધુ સારા સંકલનનું સૂચક મેટાબોલિક ફેરફારો ગ્લુકોરાફેનિનથી સમૃદ્ધ આહાર સાથે માનવ હસ્તક્ષેપના અભ્યાસમાં નોંધાયા છે, જે શાસ્ત્રીય Nrf2 એક્ટિવેટર સલ્ફોરાફેનનો પુરોગામી છે. 66].
મિટોકોન્ડ્રીયલ FAO ના પ્રથમ પગલા દરમિયાન, ?-કાર્બનનું પ્રો-આર હાઇડ્રોજન હાઇડ્રાઇડ તરીકે છોડે છે જે FAD કોફેક્ટરને FADH2 માં ઘટાડે છે, જે બદલામાં શ્વસન સાંકળમાં ઇલેક્ટ્રોનને ubiquinone (UbQ) માં સ્થાનાંતરિત કરે છે, આખરે ATP ઉત્પાદનમાં ફાળો આપે છે. . જ્યારે ગ્લુકોઝની ગેરહાજરીમાં palmitoylcarnitine દ્વારા FAO ની ઉત્તેજના WT અને Keap1-KO કોષોમાં ATP સ્તરોમાં અપેક્ષિત વધારોનું કારણ બને છે, જ્યારે Keap1-KO કોષોમાં ATP ઝડપથી વધે છે, સમાન સારવાર Nrf2-KO માં ATP ફેરફારો પેદા કરતી નથી. MEFs [65]. આ પ્રયોગ દર્શાવે છે કે, Nrf2 ની ગેરહાજરીમાં, FAO ને દબાવવામાં આવે છે, અને વધુમાં, તે Nrf2 ની ઉણપ [35], [64] ની પરિસ્થિતિમાં નીચા ATP સ્તરના એક કારણ તરીકે FAO ના દમનને સૂચિત કરે છે.
નોંધનીય રીતે, માનવીય 293 T કોષો કે જેમાં Nrf2ને શાંત કરવામાં આવ્યા છે તેમાં CPT1 અને CPT2[67] ની નીચી અભિવ્યક્તિ છે, કાર્નેટીન પાલ્મિટોયલટ્રાન્સફેરેસ (CPT) ના બે આઇસોફોર્મ છે, જે મિટોકોન્ડ્રીયલ FAO માં દર-મર્યાદિત એન્ઝાઇમ છે. કરારમાં, WT ઉંદર [1] ની સરખામણીમાં Nrf2-KO ના યકૃતમાં Cpt68 નું mRNA સ્તર ઓછું છે. સીપીટી કોએનઝાઇમ A થી l-કાર્નેટીન માં લાંબા-સાંકળ ફેટી એસિલ-CoA ના એસિલ જૂથના સ્થાનાંતરણને ઉત્પ્રેરિત કરે છે અને આ રીતે સાયટોપ્લાઝમમાંથી મિટોકોન્ડ્રિયામાં એસિલકાર્નેટીન આયાત કરવાની મંજૂરી આપે છે. જો કે આજ સુધી આની તપાસ કરવામાં આવી નથી, તે શક્ય છે કે CPT1 અભિવ્યક્તિ પર ટ્રાન્સક્રિપ્શનલ અસરો ઉપરાંત, Nrf2 તેના મુખ્ય એલોસ્ટેરિક અવરોધક, મેલોનીલ-CoA ના સ્તરને નિયંત્રિત કરીને આ એન્ઝાઇમના કાર્યને પણ અસર કરી શકે છે. આનું કારણ એ છે કે, હાલમાં અસ્પષ્ટ હોય તેવી પદ્ધતિ દ્વારા, Nrf2 સ્ટીરોયલ CoA ડેસેટ્યુરેઝ (SCD) [69] અને સાઇટ્રેટ lyase (CL) [69], [70] ની અભિવ્યક્તિને નકારાત્મક રીતે નિયંત્રિત કરે છે. વિચિત્ર રીતે, SCD નો નોકઆઉટ અથવા નિષેધ એએમપી-સક્રિય પ્રોટીન કિનેઝ (AMPK) [71], [72], [73] ના ફોસ્ફોરાયલેશન અને સક્રિયકરણ તરફ દોરી જાય છે, અને એવું અનુમાન કરી શકાય છે કે, Nrf2 ની ગેરહાજરીમાં, SCD સ્તરો વધશે, બદલામાં AMPK પ્રવૃત્તિ ઘટશે. Nrf2-KO ઉંદર [68] ના યકૃતમાં જોવા મળેલા AMPK ના ઘટેલા પ્રોટીન સ્તરો દ્વારા આને વધુ જટિલ બનાવી શકાય છે, જે કેપ1-KD ના યકૃતમાં નોંધાયેલા AMPK સ્તરો સાથે નજીકના કરારમાં હોવાનું એક તારણ છે. ઉંદર [74]. એએમપીકે પ્રવૃત્તિમાં ઘટાડો થવાનું એક પરિણામ એસીટીલ-કોએ કાર્બોક્સિલેઝ (એસીસી) [79] ના તેના અવરોધક ફોસ્ફોરાયલેશન (સેર75 પર) ની રાહત છે, જે Nrf2 ની ગેરહાજરીમાં વધુ ટ્રાન્સક્રિપ્શનલી અપરેગ્યુલેટ થઈ શકે છે કારણ કે તે Nrf2 સક્રિયકરણ દ્વારા નિયંત્રિત છે [70] ]. ઉચ્ચ ACC પ્રવૃત્તિ, અપરેગ્યુલેટેડ CL અભિવ્યક્તિ સાથે સંયોજનમાં જે એસિટિલ-CoA ના ઉત્પાદનમાં વધારો કરશે, ACC માટે સબસ્ટ્રેટ, આખરે ACC ઉત્પાદન, malonyl-CoA ના સ્તરમાં વધારો કરી શકે છે. malonyl-CoA નું ઉચ્ચ સ્તર સીપીટીને અટકાવશે, જેનાથી મિટોકોન્ડ્રિયામાં ફેટી એસિડનું પરિવહન ઘટશે. છેલ્લે, Nrf2 હકારાત્મક રીતે CD36 [76] ની અભિવ્યક્તિનું નિયમન કરે છે, એક ટ્રાન્સલોકેસ જે પ્લાઝ્મા અને મિટોકોન્ડ્રીયલ પટલમાં ફેટી એસિડની આયાત કરે છે. આમ, એક પદ્ધતિ કે જેના દ્વારા Nrf2 મિટોકોન્ડ્રીયલ FAO ની કાર્યક્ષમતાને અસર કરી શકે છે તે છે મિટોકોન્ડ્રીઆમાં લાંબા-ચેઈન ફેટી એસિડની આયાતને નિયંત્રિત કરીને.
ડાયરેક્ટ ટ્રાન્સક્રિપ્શનલ રેગ્યુલેશન ઉપરાંત, Nrf2 સેલ્યુલર રેડોક્સ મેટાબોલિઝમ પર તેની અસરો દ્વારા મિટોકોન્ડ્રીયલ FAO ની કાર્યક્ષમતામાં પણ ફેરફાર કરી શકે છે. આ ખાસ કરીને સંબંધિત હોઈ શકે છે જ્યારે Nrf2 પ્રવૃત્તિ ઓછી હોય અથવા ગેરહાજર હોય, એવી પરિસ્થિતિઓ કે જે સેલ્યુલર રેડોક્સ સ્થિતિને ઓક્સિડાઇઝ્ડ સ્થિતિ તરફ ખસેડે છે. ખરેખર, કેટલાક FAO ઉત્સેચકોને રેડોક્સ ફેરફારો પ્રત્યે સંવેદનશીલ તરીકે ઓળખવામાં આવ્યા છે. આવું એક એન્ઝાઇમ ખૂબ જ લાંબી સાંકળ એસિલ-CoA ડિહાઇડ્રોજેનેઝ (VLCAD) છે, જે માનવ પેશીઓ [80] માં palmitoyl-CoA ડિહાઇડ્રોજનેશન પ્રવૃત્તિમાં 77% થી વધુ યોગદાન આપે છે. રસપ્રદ રીતે, હર્ડ એટ અલ. [૭૮] દર્શાવે છે કે વીએલસીએડીમાં સિસ્ટીન અવશેષો હોય છે જે એકલા ઉંદરના હૃદયના મિટોકોન્ડ્રિયાના H78O2 સાથે સંપર્કમાં આવવા પર તેમની રેડોક્સ સ્થિતિમાં નોંધપાત્ર ફેરફાર કરે છે. વધુમાં, Cys2 ખાતે મ્યુરિન હેપેટિક VLCAD નું S-નાઈટ્રોસિલેશન એન્ઝાઇમ [238] ની ઉત્પ્રેરક કાર્યક્ષમતામાં સુધારો કરે છે, અને તે સંભવ છે કે સમાન સિસ્ટીનનું ઓક્સિડેશન વિપરીત અસર કરી શકે છે, જે આખરે મિટોકોન્ડ્રીયલ FAO ની કાર્યક્ષમતા ઘટાડે છે. તેથી શક્ય છે કે, WT, Nrf79-KO, અથવા Keap2-KO MEFs [1] માં VLCAD ના અભિવ્યક્તિ સ્તરો નોંધપાત્ર રીતે અલગ ન હોવા છતાં, VLCAD ની એન્ઝાઇમ પ્રવૃત્તિ ઉચ્ચ સ્તરોને કારણે Nrf65 ની ગેરહાજરીમાં ઓછી હોઈ શકે છે. ROS ના.
આ તમામ તારણોના આધારે, તે પ્રસ્તાવિત કરી શકાય છે કે (ફિગ. 3): Nrf2 ની ગેરહાજરીમાં, ME1, IDH1, G6PD અને PGD ની અભિવ્યક્તિમાં ઘટાડો થવાને કારણે NADPH સ્તરો નીચા છે. તેના જૈવસંશ્લેષણ અને પુનર્જીવનમાં ભાગ લેનારા ઉત્સેચકોની અભિવ્યક્તિમાં ઘટાડો અને ગ્લુટાથિઓનના ઘટેલા સ્વરૂપમાં ઓક્સિડાઇઝ્ડ રૂપાંતર માટે જરૂરી એનએડીપીએચના નીચલા સ્તરને કારણે ઘટાડેલા ગ્લુટાથિઓનનું સ્તર પણ ઓછું છે. ME1 ની નીચી અભિવ્યક્તિ મિટોકોન્ડ્રિયામાં પ્રવેશતા પાયરુવેટના પૂલને ઘટાડશે, જેમાં ગ્લાયકોલિસિસ પાયરુવેટનો મુખ્ય સ્ત્રોત બનશે. NADH ની પેઢી ધીમી છે, જે જટિલ I ની ક્ષતિગ્રસ્ત પ્રવૃત્તિ તરફ દોરી જાય છે અને મિટોકોન્ડ્રીયલ ROS ઉત્પાદનમાં વધારો કરે છે. FAD નું FADH2 માં ઘટાડો પણ ધીમો છે, ઓછામાં ઓછા એક ભાગમાં ઓછા કાર્યક્ષમ ફેટી એસિડ ઓક્સિડેશનને કારણે, FADH2 થી UbQ અને જટિલ III માં ઇલેક્ટ્રોન પ્રવાહ સાથે સમાધાન કરે છે. UbQH2 એ સસીનેટ ડીહાઈડ્રોજેનેઝ [80] નું સક્રિયકર્તા હોવાથી, તેની રચના ધીમી કરવાથી સસીનેટ ડીહાઈડ્રોજેનેઝની એન્ઝાઇમ પ્રવૃત્તિ ઘટી શકે છે. સુપરઓક્સાઇડ અને હાઇડ્રોજન પેરોક્સાઇડના વધેલા સ્તરો જટિલ II પ્રવૃત્તિને વધુ અટકાવી શકે છે [81]. ફેટી એસિડ ઓક્સિડેશનની ઓછી કાર્યક્ષમતા ઓક્સિડેટીવ ફોસ્ફોરીલેશનમાં મિટોકોન્ડ્રીયલ શ્વસન અને ATP ઉત્પાદન માટે સબસ્ટ્રેટની ઉપલબ્ધતામાં ઘટાડો કરવામાં ફાળો આપે છે. વળતરની પદ્ધતિ તરીકે, ગ્લાયકોલિસિસને ઉન્નત કરવામાં આવે છે. એટીપી સિન્થેઝ ??એમ જાળવવાના પ્રયાસમાં, એટીપીઝ તરીકે, વિપરીત રીતે કાર્ય કરે છે.
Nrf2 અને મિટોકોન્ડ્રીયલ બાયોજેનેસિસ
એવું નોંધવામાં આવ્યું છે કે, WT ની તુલનામાં, Nrf2-KO ઉંદરના યકૃતમાં માઇટોકોન્ડ્રીયલ સામગ્રી ઓછી હોય છે (જેમ કે પરમાણુ ડીએનએ અને મિટોકોન્ડ્રીયલના ગુણોત્તર દ્વારા નક્કી થાય છે); WT અને Nrf24-KO બંને ઉંદરોમાં 2-કલાકના ઝડપી દ્વારા આમાં વધુ ઘટાડો થાય છે; તેનાથી વિપરીત, સામાન્ય ખોરાકની સ્થિતિમાં WT થી અલગ ન હોવા છતાં, ઉચ્ચ Nrf2 પ્રવૃત્તિ ધરાવતા ઉંદરમાં મિટોકોન્ડ્રીયલ સામગ્રી ઉપવાસ [82] દ્વારા પ્રભાવિત થતી નથી. રસપ્રદ વાત એ છે કે, Nrf2 એક્ટિવેટર (R)-?-લિપોઇક એસિડ [83], [84], [85] સાથે પૂરક 3T3-L1 એડિપોસાઇટ્સ [86] માં મિટોકોન્ડ્રીયલ બાયોજેનેસિસને પ્રોત્સાહન આપે છે. ન્યુક્લિયર ટ્રાન્સક્રિપ્શનલ રેગ્યુલેટરના બે વર્ગો મિટોકોન્ડ્રીયલ બાયોજેનેસિસમાં નિર્ણાયક ભૂમિકા ભજવે છે. પ્રથમ વર્ગ ટ્રાન્સક્રિપ્શન પરિબળો છે, જેમ કે ન્યુક્લિયર રેસ્પિરેટરી ફેક્ટર 11 અને 2, જે પાંચ શ્વસન સંકુલના જનીનો એન્કોડિંગ સબ્યુનિટ્સ, મિટોકોન્ડ્રીયલ ટ્રાન્સલેશનલ ઘટકો અને હેમ બાયોસિન્થેટિક એન્ઝાઇમની અભિવ્યક્તિને નિયંત્રિત કરે છે જે મિટોકોન્ડ્રીયલ મેટ્રિક્સ [88] માં સ્થાનીકૃત છે. પિયાન્તાડોસી એટ અલ. [૮૯] એ દર્શાવ્યું છે કે ન્યુક્લિયર રેસ્પિરેટરી ફેક્ટર 89નું Nrf2-આશ્રિત ટ્રાન્સક્રિપ્શનલ અપરેગ્યુલેશન માઇટોકોન્ડ્રીયલ બાયોજેનેસિસને પ્રોત્સાહન આપે છે અને કાર્ડિયોટોક્સિક એન્થ્રાસાઇક્લાઇન કેમોથેરાપ્યુટિક એજન્ટ ડોક્સોરુબિસિનની સાયટોટોક્સિસિટી સામે રક્ષણ આપે છે. તેનાથી વિપરીત, ઝાંગ એટ અલ. [૮૨] એ નોંધ્યું છે કે Nrf1 નું આનુવંશિક સક્રિયકરણ મ્યુરિન યકૃતમાં પરમાણુ શ્વસન પરિબળ 82 ના મૂળભૂત mRNA અભિવ્યક્તિને અસર કરતું નથી.
મિટોકોન્ડ્રીયલ બાયોજેનેસિસમાં નિર્ણાયક કાર્યો સાથે ન્યુક્લિયર ટ્રાન્સક્રિપ્શનલ રેગ્યુલેટર્સનો બીજો વર્ગ ટ્રાન્સક્રિપ્શનલ કોએક્ટિવેટર્સ છે, જેમ કે પેરોક્સિસોમ પ્રોલિફેરેટર-એક્ટિવેટેડ રીસેપ્ટર? કોએક્ટિવેટર્સ (PGC)1? અને 1?, જે ટ્રાંસ્ક્રિપ્શન પરિબળો, મૂળભૂત ટ્રાન્સક્રિપ્શનલ અને આરએનએ-સ્પ્લિસિંગ મશીનરી અને હિસ્ટોન-સંશોધિત ઉત્સેચકો સાથે ક્રિયાપ્રતિક્રિયા કરે છે [88], [90], [91]. કોએક્ટિવેટર્સના PGC1 પરિવારની અભિવ્યક્તિ અસંખ્ય પર્યાવરણીય સંકેતોથી પ્રભાવિત છે. Nrf2 એક્ટિવેટર સલ્ફોરાફેન સાથે માનવ ફાઇબ્રોબ્લાસ્ટની સારવારથી માઇટોકોન્ડ્રીયલ માસમાં વધારો થાય છે અને PGC1 ના ઇન્ડક્શન થાય છે? અને PGC1? [૯૨], જોકે આ અભ્યાસમાં Nrf92 પર સંભવિત અવલંબનની તપાસ કરવામાં આવી ન હતી. જો કે, ડાયાબિટીક ઉંદર કે જેમાં Nrf2 કાં તો Keap2 જનીન હાયપોમોર્ફિક નોકડાઉન (db/db:Keap1flox/?:Nrf1+/+) દ્વારા સક્રિય થાય છે અથવા વિક્ષેપિત (db/db:Keap2flox/?:Nrf1?/?) નીચા યકૃત PGC2 ધરાવે છે? નિયંત્રણ પ્રાણીઓ કરતાં અભિવ્યક્તિ સ્તર (db/db:Keap1flox/+:Nrf1+/+) [2]. PGC93 માટે mRNA સ્તરોમાં કોઈ તફાવત નથી? નોનડાયાબિટીક ઉંદરના યકૃતમાં જોવા મળે છે જે કાં તો WT અથવા Nrf1-KO છે, જ્યારે Nrf2-ઓવરએક્સપ્રેસિંગ (Keap2-KD અને લીવર-વિશિષ્ટ Keap1-KO) પ્રાણીઓમાં આ સ્તર નીચું છે [1]. નોંધનીય છે કે, 82-કલાકનો ઉપવાસ PGC24 નું સ્તર વધારે છે? તમામ જીનોટાઇપના ઉંદરના યકૃતમાં mRNA, પરંતુ WT અથવા Nrf1-ઓવરએક્સપ્રેસિંગ ઉંદરોની સરખામણીમાં Nrf2-KO ના યકૃતમાં નોંધપાત્ર વધારો છે. WT ની તુલનામાં, Nrf2-KO ઉંદર ચેપને કારણે સેપ્ટિક ચેપ અથવા તીવ્ર ફેફસાની ઇજાનો અનુભવ કરે છે તે પરમાણુ શ્વસન પરિબળ 2 અને PGC1 નું એટેન્યુએટેડ ટ્રાન્સક્રિપ્શનલ અપગ્ર્યુલેશન દર્શાવે છે? [1], [94]. એકસાથે, આ અવલોકનો સૂચવે છે કે પરમાણુ શ્વસન પરિબળ 95 અને PGC2 બંનેના સ્તરને જાળવવામાં Nrf1 ની ભૂમિકા? જટિલ છે અને તણાવની પરિસ્થિતિઓમાં સૌથી વધુ અગ્રણી બને છે.
મિટોકોન્ડ્રીયલ પ્રોટીનના એન્કોડિંગ જનીનોની અભિવ્યક્તિ ઉપરાંત, મિટોકોન્ડ્રીયલ બાયોજેનેસિસ માટે ન્યુક્લિયોટાઇડ્સના સંશ્લેષણની જરૂર છે. Nrf2 નું આનુવંશિક સક્રિયકરણ પેન્ટોઝ ફોસ્ફેટ પાથવે અને ફોલેટ અને ગ્લુટામાઈનના ચયાપચયને અપરેગ્યુલેટ કરીને પ્યુરિન બાયોસિન્થેસિસને વધારે છે, ખાસ કરીને ઝડપથી ફેલાતા કોષોમાં (ફિગ. 2) [24]. મિટોકોન્ડ્રીયલ સેરીન/થ્રેઓનિન પ્રોટીન કિનેઝ PTEN-પ્રેરિત પુટેટિવ કિનેઝ 1 (PINK1) માટે મ્યુટન્ટ ડ્રોસોફિલાની ઉણપના ટ્રાન્સક્રિપ્ટોમનું વિશ્લેષણ દર્શાવે છે કે માઇટોકોન્ડ્રીયલ ડિસફંક્શન જનીનોના ટ્રાન્સક્રિપ્શનલ અપગ્ર્યુલેશન તરફ દોરી જાય છે જે ન્યુક્લિયોટાઇડ્સ અને બાયોટોટાઇડને અસર કરે છે. PINK96 ની ઉણપના ન્યુરોટોક્સિક પરિણામો સામે રક્ષણ માટેની પદ્ધતિ રજૂ કરે છે. Nrf1 ફોસ્ફોરીબોસિલ પાયરોફોસ્ફેટ એમીડોટ્રાન્સફેરેઝ (PPAT) ની અભિવ્યક્તિનું નિયમન કરે છે, જે ડી નોવો પ્યુરિન ન્યુક્લિયોટાઈડ બાયોસિન્થેટિક પાથવે અને મિટોકોન્ડ્રીયલ મેથિલેનેટેટ્રાહાઈડ્રોફોલેટ ડીહાઈડ્રોજેનેઝ 2 (MTHigFD2) માં પ્રવેશને ઉત્પ્રેરિત કરે છે. બાદમાં ડીહાઇડ્રોજેનેઝ અને સાયક્લોહાઇડ્રોલેઝ પ્રવૃત્તિઓ સાથેનું દ્વિ-કાર્યકારી એન્ઝાઇમ છે જે ઝડપથી વિકસતા કોષોમાં પ્યુરિન બાયોસિન્થેસિસ માટે એક-કાર્બન એકમોના સ્ત્રોત તરીકે ગ્લાયસીન અને ફોર્મેટ બંને પ્રદાન કરવામાં મહત્વપૂર્ણ છે [2]. તેથી સંભવ છે કે Nrf2 સક્રિયકરણ રક્ષણાત્મક હોઈ શકે છે અને PINK97 ની ઉણપમાં મિટોકોન્ડ્રીયલ ડિસફંક્શનને ઉલટાવી શકે છે. ખરેખર, સલ્ફોરાફેન દ્વારા Nrf2 નું ફાર્માકોલોજીકલ સક્રિયકરણ, અથવા ટ્રાઇટરપેનોઇડ RTA-1, ??m પુનઃસ્થાપિત કરે છે અને ડોપામાઇન ટોક્સિસિટી [2] સામે PINK408-ઉણપવાળા કોષોનું રક્ષણ કરે છે. જો કે અંતર્ગત પદ્ધતિઓ જટિલ લાગે છે, એકસાથે, આ તારણો સૂચવે છે કે Nrf1 પ્રવૃત્તિ નિર્ણાયક ટ્રાન્સક્રિપ્શન પરિબળો અને કોએક્ટિવેટર્સના અભિવ્યક્તિ સ્તરોને પ્રભાવિત કરીને તેમજ ન્યુક્લિયોટાઇડ બાયોસિન્થેસિસને વધારીને મિટોકોન્ડ્રીયલ બાયોજેનેસિસને અસર કરી શકે છે.
Nrf2 અને મિટોકોન્ડ્રીયલ અખંડિતતા
જોકે પ્રત્યક્ષ પુરાવા હંમેશા ઉપલબ્ધ હોતા નથી, એવા મજબૂત સંકેતો છે કે Nrf2 એ મિટોકોન્ડ્રીયલ અખંડિતતા માટે મહત્વપૂર્ણ છે, ખાસ કરીને ઓક્સિડેટીવ તણાવની સ્થિતિમાં. Nrf2 એક્ટિવેટર સલ્ફોરાફેનની એક માત્રા આપવામાં આવી હોય તેવા ઉંદરોના મગજ અને યકૃતમાંથી અલગ પડેલ મિટોકોન્ડ્રિયા ઓક્સિડન્ટ ટર્ટ-બ્યુટિલહાઇડ્રોપેરોક્સાઇડ [99], [100] ને કારણે મિટોકોન્ડ્રીયલ અભેદ્યતા સંક્રમણ છિદ્ર (mPTP) ખોલવા માટે પ્રતિરોધક છે. એમપીટીપી, એક સંકુલ કે જે મિટોકોન્ડ્રીયલ આંતરિક પટલને 1500 Da સુધીના સમૂહ સાથેના અણુઓ માટે અભેદ્ય બનવા માટે પરવાનગી આપે છે, તે તાજેતરમાં F0F1-ATP સિન્થેઝ [101] ના ડાઇમર્સમાંથી રચાયેલ હોવાનું જાણવા મળ્યું હતું. એમપીટીપી ખોલવા માટે સલ્ફોરાફેન-મધ્યસ્થી પ્રતિકાર વધેલા એન્ટીઑકિસડન્ટ સંરક્ષણ સાથે સંબંધ ધરાવે છે, અને માઇટોકોન્ડ્રીયલ જીએસએચ, ગ્લુટાથિઓન પેરોક્સિડેઝ 1, મેલિક એન્ઝાઇમ 3, અને થિયોરેડોક્સિન 2 ના સ્તરો તમામ માઇટોકોન્ડ્રીયલ અપૂર્ણાંકમાં અપરેગ્યુલેટેડ છે જે પ્રાણીઓ [ટ્રેલ્ફોરાફેન] 100 થી અલગ છે.
ઇલેક્ટ્રોફિલિક લિપિડ પેરોક્સિડેશન પ્રોડક્ટ 4-હાઇડ્રોક્સી-2-નોનેનલને કારણે મિટોકોન્ડ્રીયલ પ્રોટીનનું નુકસાન અને શ્વસનમાં ક્ષતિ, સલ્ફોરાફેન-સારવાર કરાયેલ ઉંદરના સેરેબ્રલ કોર્ટેક્સ [102] થી અલગ કરાયેલા મિટોકોન્ડ્રિયામાં ક્ષીણ થાય છે. ઉંદરના મૂત્રપિંડના ઉપકલા કોષોમાં અને કિડનીમાં, સલ્ફોરાફેન સિસ્પ્લેટિન- અને જેન્ટામિસિન-પ્રેરિત ઝેરી અને ??m[103], [104] ના નુકશાન સામે રક્ષણાત્મક છે. ઓક્સિડન્ટ્સ (સુપરઓક્સાઇડ, હાઇડ્રોજન પેરોક્સાઇડ, પેરોક્સાઇનાઇટ) અને ઇલેક્ટ્રોફાઇલ્સ (4-હાઇડ્રોક્સી-2-નોનેનલ અને એક્રોલિન) સામે રક્ષણ અને સલ્ફોરાફેન [105] સાથે ઉંદરની એઓર્ટિક સ્મૂથ સ્નાયુ કોશિકાઓની સારવાર પર મિટોકોન્ડ્રીયલ એન્ટીઑકિસડન્ટ સંરક્ષણમાં વધારો પણ જોવા મળ્યો છે. ]. કોન્ટ્રાસ્ટ-પ્રેરિત તીવ્ર કિડનીની ઇજાના મોડેલમાં, અંગની ઇસ્કેમિક પૂર્વશરત તાજેતરમાં GSK2 ના નિષેધને પરિણામે Nrf3 ના સક્રિયકરણ દ્વારા, એમપીટીપી અને મિટોકોન્ડ્રીયલ સોજોના ઉદઘાટનના નિષેધ સહિત રક્ષણાત્મક અસરો દર્શાવવામાં આવી હતી? [106].
મિટોફેજી, પ્રક્રિયા કે જેના દ્વારા નિષ્ક્રિય મિટોકોન્ડ્રિયા ઓટોફેગોસોમ્સ દ્વારા પસંદગીયુક્ત રીતે આવરી લેવામાં આવે છે અને કોષ દ્વારા ડિગ્રેડ અને રિસાયકલ કરવા માટે લાઇસોસોમ સુધી પહોંચાડવામાં આવે છે, તે મિટોકોન્ડ્રીયલ હોમિયોસ્ટેસિસ [107], [108] માટે આવશ્યક છે. જ્યારે Nrf2 અને મિટોફેજી વચ્ચે કોઈ કારણભૂત સંબંધ સ્થાપિત થયો નથી, ત્યાં પુરાવા છે કે મિટોફેજીમાં ભૂમિકા ભજવીને માઇટોકોન્ડ્રીયલ ગુણવત્તા નિયંત્રણમાં ટ્રાન્સક્રિપ્શન પરિબળ મહત્વપૂર્ણ હોઈ શકે છે. ઓક્સિડેટીવ તણાવની પરિસ્થિતિઓમાં આ ખાસ કરીને અગ્રણી હોઈ શકે છે. આમ, સેપ્સિસના મોડેલમાં, WT ઉંદરની સરખામણીમાં Nrf1-KO માં ઓટોફેગોસોમ માર્કર MAP3 લાઇટ ચેઇન 3-II (LC62-II) અને કાર્ગો પ્રોટીન p24 ના સ્તરમાં 2 કલાક પછી વધારો દબાવવામાં આવે છે [109] . મિટોફેજીનું એક નાનું-પરમાણુ પ્રેરક (જેને p62-મીડિયેટેડ મિટોફેજી ઇન્ડ્યુસર, PMI કહેવાય છે) તાજેતરમાં શોધાયું હતું; આ 1,4-ડિફેનાઇલ-1,2,3-ટ્રાઇઝોલ સંયોજન મૂળરૂપે Nrf2 એક્ટિવેટર તરીકે ડિઝાઇન કરવામાં આવ્યું હતું જે કેપ1 [110] સાથે ટ્રાન્સક્રિપ્શન પરિબળની ક્રિયાપ્રતિક્રિયાને અવરોધે છે. કોષોની જેમ કે જેમાં Nrf2 આનુવંશિક રીતે અપરેગ્યુલેટેડ છે (Keap1-KD અથવા Keap1-KO), PMIના સંપર્કમાં આવેલા કોષો વધુ આરામ કરે છે ??m. અગત્યની રીતે, માઇટોકોન્ડ્રીયલ LC3 સ્થાનિકીકરણમાં વધારો જે WT કોષોની PMI સારવાર પછી જોવા મળે છે તે Nrf2-KO કોષોમાં થતો નથી, જે Nrf2 ની સંડોવણી સૂચવે છે.
છેલ્લે, યકૃતના વિભાગોના અલ્ટ્રાસ્ટ્રક્ચરલ પૃથ્થકરણે Nrf2-KO ના હિપેટોસાઇટ્સમાં ઘટેલા ક્રિસ્ટા અને વિક્ષેપિત પટલ સાથેના સોજાવાળા મિટોકોન્ડ્રિયાની હાજરી જાહેર કરી છે, પરંતુ WT નહીં, ઉંદરો કે જેમને 24 અઠવાડિયાથી વધુ ચરબીયુક્ત ખોરાક આપવામાં આવ્યો હતો; ખાસ કરીને, આ યકૃત ઓક્સિડેટીવ તણાવ અને બળતરાના સ્પષ્ટ પુરાવા દર્શાવે છે [68]. તે નિષ્કર્ષ પર આવી શકે છે કે ઓક્સિડેટીવ અને બળતરા તણાવની પરિસ્થિતિઓમાં માઇટોકોન્ડ્રીયલ અખંડિતતા જાળવવામાં Nrf2 ની મહત્વપૂર્ણ ભૂમિકા છે.
સલ્ફોરાફેન અને કેન્સર, મૃત્યુદર, વૃદ્ધત્વ, મગજ અને વર્તન, હૃદય રોગ અને વધુ પર તેની અસરો
Isothiocyanates એ કેટલાક સૌથી મહત્વપૂર્ણ છોડ સંયોજનો છે જે તમે તમારા આહારમાં મેળવી શકો છો. આ વિડિયોમાં હું તેમના માટે અત્યાર સુધીનો સૌથી વ્યાપક કેસ બનાવું છું. ટૂંકા ધ્યાન ગાળો? નીચેના ટાઈમ પોઈન્ટ્સમાંથી કોઈ એક પર ક્લિક કરીને તમારા મનપસંદ વિષય પર જાઓ. નીચે સંપૂર્ણ સમયરેખા.
મુખ્ય વિભાગો:
00:01:14 - કેન્સર અને મૃત્યુદર
00:19:04 - વૃદ્ધત્વ
00:26:30 - મગજ અને વર્તન
00:38:06 - અંતિમ રીકેપ
00:40:27 - માત્રા
સંપૂર્ણ સમયરેખા:
00:00:34 - સલ્ફોરાફેનનો પરિચય, વિડીયોનું મુખ્ય ધ્યાન.
00:01:14 - ક્રુસિફેરસ શાકભાજીનો વપરાશ અને સર્વ-કારણ મૃત્યુદરમાં ઘટાડો.
00:44:24 - શાકભાજીમાંથી સક્રિય માયરોસિનેઝ સાથે જોડવામાં આવે ત્યારે પૂરક વધુ સારી રીતે કામ કરે છે.
00:44:56 - રાંધવાની તકનીકો અને ક્રુસિફેરસ શાકભાજી.
00:46:06 - ગોઇટ્રોજન તરીકે આઇસોથિયોસાયનેટ્સ.
Nrf2 એ એક ટ્રાન્સક્રિપ્શન પરિબળ છે જે માનવ શરીરની સેલ્યુલર એન્ટીઑકિસડન્ટ સંરક્ષણ પ્રણાલીમાં મહત્વપૂર્ણ ભૂમિકા ભજવે છે. એન્ટીઑકિસડન્ટ પ્રતિભાવ તત્વ, અથવા ARE, જનીનોની એક નિયમનકારી પદ્ધતિ છે. ઘણા સંશોધન અભ્યાસોએ દર્શાવ્યું છે કે Nrf2, અથવા NF-E2-સંબંધિત પરિબળ 2, વિવિધ પ્રકારના કોષોમાં ARE-સંચાલિત જનીનોનું નિયમન કરે છે. Nrf2 એ સેલ્યુલર પ્રોટેક્શન અને એન્ટિ-કાર્સિનોજેનિસિટીમાં પણ આવશ્યક ભૂમિકા ભજવી હોવાનું જણાયું હતું, જે દર્શાવે છે કે Nrf2 એ ઓક્સિડેટીવ તણાવને કારણે માનવામાં આવતા ન્યુરોડિજનરેટિવ રોગો અને કેન્સરના સંચાલનમાં અસરકારક સારવાર હોઈ શકે છે. ડૉ. એલેક્સ જિમેનેઝ ડીસી, CCST ઇનસાઇટ
સમાપન નોંધ, ઉપસંહાર
જો કે ઘણા પ્રશ્નો હજુ પણ ખુલ્લા છે, ઉપલબ્ધ પ્રાયોગિક પુરાવા સ્પષ્ટપણે સૂચવે છે કે Nrf2 એ મિટોકોન્ડ્રીયલ હોમિયોસ્ટેસિસ અને માળખાકીય અખંડિતતાની જાળવણીમાં એક મહત્વપૂર્ણ ખેલાડી છે. આ ભૂમિકા ઓક્સિડેટીવ, ઇલેક્ટ્રોફિલિક અને દાહક તાણની પરિસ્થિતિઓમાં ખાસ કરીને નિર્ણાયક બની જાય છે જ્યારે Nrf2- મધ્યસ્થી સાયટોપ્રોટેક્ટીવ પ્રતિભાવોને અપગ્રેજ્યુલેટ કરવાની ક્ષમતા કોષ અને જીવતંત્રના એકંદર આરોગ્ય અને અસ્તિત્વને પ્રભાવિત કરે છે. માઇટોકોન્ડ્રીયલ કાર્યમાં Nrf2 ની ભૂમિકા આ ટ્રાન્સક્રિપ્શન પરિબળ દ્વારા ગોઠવવામાં આવેલ વ્યાપક સાયટોપ્રોટેક્ટીવ મિકેનિઝમ્સના બીજા સ્તરને રજૂ કરે છે. ઘણા માનવ રોગવિજ્ઞાનની પરિસ્થિતિઓમાં ઓક્સિડેટીવ તણાવ, બળતરા અને માઇટોકોન્ડ્રીયલ ડિસફંક્શન તેમના પેથોજેનેસિસના આવશ્યક ઘટકો તરીકે હોય છે, Nrf2 નું ફાર્માકોલોજિકલ સક્રિયકરણ રોગ નિવારણ અને સારવાર માટે વચન ધરાવે છે. ચોક્કસ મિકેનિઝમ્સની વ્યાપક સમજ કે જેના દ્વારા Nrf2 મિટોકોન્ડ્રીયલ કાર્યને અસર કરે છે તે ભવિષ્યના ક્લિનિકલ ટ્રાયલ્સની તર્કસંગત રચના માટે જરૂરી છે અને રોગનિવારક અસરકારકતાનું નિરીક્ષણ કરવા માટે નવા બાયોમાર્કર્સ ઓફર કરી શકે છે.
ઉપરોક્ત લેખનો હેતુ મિટોકોન્ડ્રીયલ કાર્યમાં Nrf2 ની ઉભરતી ભૂમિકાની ચર્ચા કરવાનો તેમજ દર્શાવવાનો હતો. Nrf2, અથવા પરમાણુ પરિબળ એરિથ્રોઇડ 2-સંબંધિત પરિબળ, ઓક્સિડન્ટ્સ માટે સેલ્યુલર પ્રતિકારનું ઉભરતું નિયમનકાર છે જે ઓક્સિડેટીવ તણાવમાં ફાળો આપી શકે છે, સેલ્યુલર કાર્યને અસર કરે છે અને ઝેરી, ક્રોનિક રોગ અને કેન્સરના વિકાસ તરફ દોરી જાય છે. જ્યારે માનવ શરીરમાં ઓક્સિડન્ટ્સનું ઉત્પાદન કોષ વિભાજન, બળતરા, રોગપ્રતિકારક કાર્ય, ઓટોફેજી અને તણાવ પ્રતિભાવ સહિત વિવિધ હેતુઓ પૂરા કરી શકે છે, ત્યારે સ્વાસ્થ્ય સમસ્યાઓને રોકવા માટે તેમના અતિશય ઉત્પાદનને નિયંત્રિત કરવું આવશ્યક છે. અમારી માહિતીનો અવકાશ ચિરોપ્રેક્ટિક અને કરોડરજ્જુના સ્વાસ્થ્ય સમસ્યાઓ સુધી મર્યાદિત છે. વિષયની ચર્ચા કરવા માટે, કૃપા કરીને ડૉ. જીમેનેઝને પૂછો અથવા અમારો સંપર્ક કરો915-850-0900 .
પીઠનો દુખાવોવિકલાંગતાના સૌથી પ્રચલિત કારણોમાંનું એક અને કામના દિવસો ચૂકી જવાના દિવસો છે. પીઠનો દુખાવો એ ડૉક્ટરની ઑફિસની મુલાકાત માટેના બીજા સૌથી સામાન્ય કારણને આભારી છે, જે ફક્ત ઉપલા-શ્વસન માર્ગના ચેપથી વધુ છે. આશરે 80 ટકા વસ્તી તેમના સમગ્ર જીવનમાં ઓછામાં ઓછા એક વખત પીઠનો દુખાવો અનુભવશે. કરોડરજ્જુ એ હાડકાં, સાંધા, અસ્થિબંધન અને સ્નાયુઓ, અન્ય નરમ પેશીઓની વચ્ચે બનેલી જટિલ રચના છે. આને કારણે, ઇજાઓ અને/અથવા વિકટ પરિસ્થિતિ, જેમ કે�હર્નિયેટ ડિસ્ક, આખરે પીઠના દુખાવાના લક્ષણો તરફ દોરી શકે છે. રમતગમતની ઇજાઓ અથવા ઓટોમોબાઇલ અકસ્માતની ઇજાઓ પીઠના દુખાવા માટેનું સૌથી વારંવારનું કારણ છે, જો કે, કેટલીકવાર સરળ હલનચલન પીડાદાયક પરિણામો લાવી શકે છે. સદનસીબે, વૈકલ્પિક સારવાર વિકલ્પો, જેમ કે ચિરોપ્રેક્ટિક સંભાળ, કરોડરજ્જુના ગોઠવણો અને મેન્યુઅલ મેનિપ્યુલેશન્સના ઉપયોગ દ્વારા પીઠનો દુખાવો ઓછો કરવામાં મદદ કરી શકે છે, આખરે પીડા રાહતમાં સુધારો કરે છે. �
એનઆરએફએક્સએનએક્સ એન્ટીઑકિસડન્ટ અને ડિટોક્સિફાઇંગ એન્ઝાઇમ્સ અને જનીનોના જૂથના સક્રિયકરણને સમર્થન આપે છે જે માનવ શરીરને ઓક્સિડેટીવ તણાવના વધેલા સ્તર સાથે સંકળાયેલ આરોગ્ય સમસ્યાઓની અસરોથી રક્ષણ આપે છે, જેમ કે અલ્ઝાઇમર રોગ. Nrf2 પાથવેને સક્રિય કરવા માટે વિવિધ પ્રકારના કુદરતી પદાર્થોનું નિદર્શન કરવામાં આવ્યું છે, જે ન્યુરોડિજનરેટિવ રોગોના લક્ષણોનું સંચાલન કરવામાં મદદ કરી શકે છે. નીચેના લેખનો હેતુ ક્રોનિક સોજાને કારણે Nrf2 ની મુખ્ય ભૂમિકાની ચર્ચા કરવાનો છે.
અમૂર્ત
કાર્સિનોજેનેસિસમાં નિર્ણાયક ભૂમિકા ભજવતી વખતે, ઘણા ક્રોનિક રોગો અને ગૂંચવણોમાં બળતરા એ સૌથી સામાન્ય લક્ષણ છે. કેટલાક અભ્યાસોએ દર્શાવ્યું છે કે Nrf2 બળતરા વિરોધી પ્રક્રિયામાં ફાળો આપે છે બળતરા કોશિકાઓની ભરતી અને એન્ટીઑકિસડન્ટ પ્રતિભાવ તત્વ (ARE) દ્વારા જનીન અભિવ્યક્તિનું નિયમન કરીને. Keap1 (Kelch-like ECH-સંબંધિત પ્રોટીન)/Nrf2 (NF-E2 p45-સંબંધિત પરિબળ 2)/ARE સિગ્નલિંગ પાથવે મુખ્યત્વે બળતરા વિરોધી જનીન અભિવ્યક્તિને નિયંત્રિત કરે છે અને બળતરાની પ્રગતિને અટકાવે છે. તેથી, નવા Nrf2-આશ્રિત બળતરા વિરોધી ફાયટોકેમિકલ્સની ઓળખ દવાની શોધમાં મુખ્ય મુદ્દો બની ગયો છે. આ સમીક્ષામાં, અમે Keap1/Nrf2/ARE સિગ્નલ પાથવેના સભ્યો અને તેના ડાઉનસ્ટ્રીમ જનીનો, દાહક રોગોના પ્રાણી મોડલ પર આ માર્ગની અસરો અને NF-?B પાથવે સાથે ક્રોસસ્ટૉકની ચર્ચા કરીએ છીએ. વધુમાં અમે Nrf3 દ્વારા NLRP2 બળતરાના નિયમન વિશે પણ ચર્ચા કરીએ છીએ. આ ઉપરાંત, અમે Nrf2/ARE સિગ્નલિંગ પાથવેને મધ્યસ્થી કરતા એન્ટિ-ઇન્ફ્લેમેટરી ફાયટોકેમિકલ્સ અને અન્યના વિકાસના વર્તમાન દૃશ્યનો સારાંશ આપીએ છીએ.
બળતરા એ એક જટિલ પ્રક્રિયા છે જે ત્યારે થાય છે જ્યારે પેથોજેન્સ, નુકસાન અથવા બળતરા જેવા હાનિકારક ઉત્તેજના દ્વારા પેશીઓને ચેપ અથવા ઇજા થાય છે. રોગપ્રતિકારક કોષો, રક્તવાહિનીઓ અને પરમાણુ મધ્યસ્થીઓ આ રક્ષણાત્મક પ્રતિભાવમાં સામેલ છે [1]. બળતરા એ એક રોગવિજ્ઞાનવિષયક ઘટના છે જે મુખ્યત્વે શારીરિક, રાસાયણિક, જૈવિક અને મનોવૈજ્ઞાનિક પરિબળો દ્વારા પ્રેરિત વિવિધ રોગની સ્થિતિઓ સાથે સંકળાયેલ છે. બળતરાનો ઉદ્દેશ્ય સેલ્યુલર નુકસાનના કારણોને મર્યાદિત અને દૂર કરવાનો છે, નેક્રોટિક કોશિકાઓ અને પેશીઓને સ્પષ્ટ અને/અથવા શોષી લે છે, અને પેશીઓની સમારકામ શરૂ કરે છે. બળતરાના બે વિશિષ્ટ સ્વરૂપોને અલગ પાડવામાં આવે છે: તીવ્ર અને ક્રોનિક. તીવ્ર બળતરા સ્વ-મર્યાદિત અને યજમાન માટે ફાયદાકારક છે, પરંતુ લાંબા સમય સુધી ક્રોનિક સોજા એ ઘણા ક્રોનિક રોગો અને ગૂંચવણોનું સામાન્ય લક્ષણ છે. ઘણા મોનોન્યુક્લિયર રોગપ્રતિકારક કોષો જેમ કે મોનોસાઇટ્સ, મેક્રોફેજેસ, લિમ્ફોસાઇટ્સ અને પ્લાઝ્મા કોશિકાઓ દ્વારા પ્રત્યક્ષ ઘૂસણખોરી, તેમજ બળતરા સાઇટોકીન્સનું ઉત્પાદન, ક્રોનિક બળતરા તરફ દોરી જાય છે. તે માન્ય છે કે ક્રોનિક સોજા કાર્સિનોજેનેસિસ [2] માં નિર્ણાયક ભૂમિકા ભજવે છે. સામાન્ય રીતે, બંને તરફી અને બળતરા વિરોધી સિગ્નલિંગ માર્ગો સામાન્ય બળતરા પ્રક્રિયામાં ક્રિયાપ્રતિક્રિયા કરે છે.
રોગવિજ્ઞાનવિષયક બળતરા પ્રક્રિયામાં, માસ્ટ કોશિકાઓ, મોનોસાઇટ્સ, મેક્રોફેજ, લિમ્ફોસાઇટ્સ અને અન્ય રોગપ્રતિકારક કોષો પ્રથમ સક્રિય થાય છે. પછી કોશિકાઓને ઇજાના સ્થળે ભરતી કરવામાં આવે છે, પરિણામે પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓ (ROS) નું નિર્માણ થાય છે જે ડીએનએ સહિત મેક્રોમોલેક્યુલ્સને નુકસાન પહોંચાડે છે. તે જ સમયે, આ બળતરા કોશિકાઓ સાયટોકાઇન્સ, કેમોકાઇન્સ અને પ્રોસ્ટાગ્લાન્ડિન્સ જેવા મોટા પ્રમાણમાં બળતરા મધ્યસ્થીઓ પણ ઉત્પન્ન કરે છે. આ મધ્યસ્થીઓ બળતરાના સ્થાનિક સ્થળોએ મેક્રોફેજની વધુ ભરતી કરે છે અને બળતરા સાથે સંકળાયેલ બહુવિધ સિગ્નલ ટ્રાન્સડક્શન કાસ્કેડ્સ અને ટ્રાન્સક્રિપ્શન પરિબળોને સીધા સક્રિય કરે છે. NF-?B (પરમાણુ પરિબળ કપ્પા B), MAPK (મિટોજન-સક્રિયકૃત પ્રોટીન કિનેઝ), અને JAK (જાનુસ કિનેઝ)-STAT (સિગ્નલ ટ્રાન્સડ્યુસર્સ અને ટ્રાન્સક્રિપ્શનના સક્રિયકર્તા) સિગ્નલ પાથવે બળતરાના શાસ્ત્રીય માર્ગના વિકાસમાં સામેલ છે. [3], [4], [5]. અગાઉના અભ્યાસોએ જાહેર કર્યું છે કે ટ્રાન્સક્રિપ્શન ફેક્ટર Nrf2 (NF-E2 p45-સંબંધિત પરિબળ 2) NADPH, NAD(P)H ક્વિનોન ઓક્સિડોરેડક્ટેઝ 1, ગ્લુટાથિઓન પેરોક્સિડેઝ, ફેરીટીન, હેમ ઓક્સિજન (HO-1) સહિત બીજા તબક્કાના ડિટોક્સિફાઇંગ એન્ઝાઇમ્સની અભિવ્યક્તિને નિયંત્રિત કરે છે. -1), અને એન્ટીઑકિસડન્ટ જનીનો જે કોષોને તેમની બળતરા વિરોધી અસરો દ્વારા વિવિધ ઇજાઓથી રક્ષણ આપે છે, આમ રોગ [6], [7], [8] ને પ્રભાવિત કરે છે.
આ નોંધપાત્ર તારણોને ધ્યાનમાં લેતા, સિગ્નલિંગ પાથવે દ્વારા બળતરા રોગો માટે લક્ષિત ઉપચારાત્મક દવાઓના વિકાસે તાજેતરના વર્ષોમાં ખૂબ જ રસ ખેંચ્યો છે. આ સમીક્ષામાં, અમે Keap1 (Kelch-like ECH સંકળાયેલ પ્રોટીન)/Nrf2 (NF-E2 p45-સંબંધિત પરિબળ 2)/ARE (એન્ટીઑકિસડન્ટ પ્રતિભાવ તત્વ) બળતરામાં સિગ્નલિંગ પાથવે પર સંશોધનનો સારાંશ આપીએ છીએ.
Nrf2 નું માળખું અને નિયમન
Keap1-આશ્રિત Nrf2 નિયમન
Nrf2 એ Cap �n� કોલર (CNC) સબફેમિલીથી સંબંધિત છે અને તેમાં સાત કાર્યાત્મક ડોમેન્સનો સમાવેશ થાય છે, નેહ (Nrf2-ECH હોમોલોજી) 1 થી Neh7 [9], [10]. Neh1 એ CNC-bZIP ડોમેન છે જે Nrf2 ને નાના મસ્ક્યુલોપોન્યુરોટિક ફાઈબ્રોસારકોમા (Maf) પ્રોટીન, DNA અને અન્ય ટ્રાન્સક્રિપ્શન ભાગીદારો સાથે હેટરોડાઈમરાઈઝ કરવા તેમજ યુબીક્વિટીન-કન્જુગેટીંગ એન્ઝાઇમ UbcM2 [11], [12] સાથે ન્યુક્લિયર કોમ્પ્લેક્સ બનાવવાની મંજૂરી આપે છે. Neh2 માં DLG અને ETGE તરીકે ઓળખાતા બે મહત્વના ઉદ્દેશો છે, જે Nrf2 અને તેના નકારાત્મક નિયમનકાર Keap1 [13], [14] વચ્ચેની ક્રિયાપ્રતિક્રિયા માટે જરૂરી છે.
Keap1 એ કુલીન-આધારિત E3 ubiquitin ligase માટે સબસ્ટ્રેટ એડેપ્ટર છે, જે સામાન્ય પરિસ્થિતિઓ [2], [15], [16] હેઠળ સર્વવ્યાપકીકરણ અને પ્રોટીસોમલ ડિગ્રેડેશન દ્વારા Nrf17 ની ટ્રાન્સક્રિપ્શનલ પ્રવૃત્તિને અટકાવે છે. Keap1 homodimer ના KELCH ડોમેન્સ cytosol માં Nrf2-Neh2 ડોમેનના DLG અને ETGE મોટિફ્સ સાથે જોડાય છે, જ્યાં ETGE ઉચ્ચ જોડાણ સાથે હિન્જ તરીકે કામ કરે છે અને DLG લૅચ તરીકે કામ કરે છે [18]. ઓક્સિડેટીવ તણાવ હેઠળ અથવા Nrf2 એક્ટિવેટર્સના સંપર્કમાં આવવા પર, Nrf2 Keap1 સિસ્ટીન અવશેષોના થિયોલ ફેરફારને કારણે Keap1 બંધનમાંથી અલગ થઈ જાય છે જે આખરે Nrf2 સર્વવ્યાપકતા અને પ્રોટીસોમલ ડિગ્રેડેશન [19] અટકાવે છે. પછી Nrf2 ન્યુક્લિયસમાં સ્થાનાંતરિત થાય છે, નાના Maf પ્રોટીન સાથે હેટરોડાઈમરાઈઝ થાય છે, અને ARE જનીનોની બેટરી (ફિગ. 1A) ને ટ્રાન્સએક્ટિવ કરે છે. Neh3 નું કાર્બોક્સી-ટર્મિનલ CHD6 (chromo-ATPase/helicase DNA બાઈન્ડિંગ પ્રોટીન) [20] તરીકે ઓળખાતા ટ્રાન્સક્રિપ્શન કો-એક્ટિવેટર સાથે ક્રિયાપ્રતિક્રિયા કરીને ટ્રાન્સએક્ટિવેશન ડોમેન તરીકે કાર્ય કરે છે. Neh4 અને Neh5 પણ ટ્રાન્સએક્ટિવેશન ડોમેન્સ તરીકે કામ કરે છે, પરંતુ CBP (cAMP-રિસ્પોન્સ-એલિમેન્ટ-બાઈન્ડિંગ પ્રોટીન-બાઈન્ડિંગ પ્રોટીન) [21] તરીકે ઓળખાતા અન્ય ટ્રાન્સક્રિપ્શનલ કો-એક્ટિવેટરને જોડે છે. વધુમાં, Neh4 અને Neh5 ન્યુક્લિયર કોફેક્ટર RAC3/AIB1/SRC-3 સાથે ક્રિયાપ્રતિક્રિયા કરે છે, જે Nrf2-લક્ષિત ARE જનીન અભિવ્યક્તિ [22] તરફ દોરી જાય છે. Neh5 પાસે રેડોક્સ-સંવેદનશીલ પરમાણુ-નિકાસ સિગ્નલ છે જે Nrf2 [23] ના નિયમન અને સેલ્યુલર સ્થાનિકીકરણ માટે નિર્ણાયક છે.
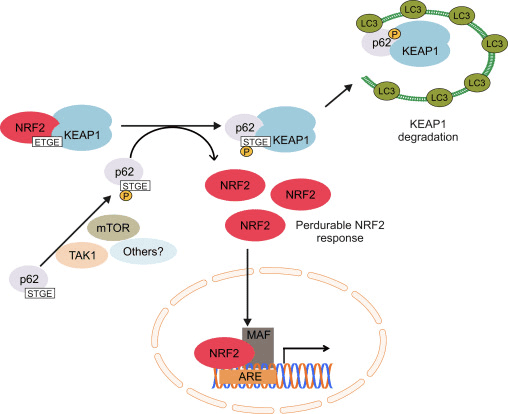
આકૃતિ 1 Nrf1 નું Keap2-આશ્રિત અને સ્વતંત્ર નિયમન. (A) મૂળભૂત પરિસ્થિતિઓ હેઠળ, Nrf2 ને તેના બે ઉદ્દેશો (ETGE અને DLG) દ્વારા Keap1 સાથે અલગ કરવામાં આવે છે જે CUL3-મધ્યસ્થી સર્વવ્યાપકતા તરફ દોરી જાય છે અને ત્યારબાદ પ્રોટીઝોમ ડિગ્રેડેશન થાય છે. ઓક્સિડેટીવ તણાવ હેઠળ, Nrf2 Keap1 થી અલગ થઈ જાય છે, ન્યુક્લિયસમાં સ્થાનાંતરિત થાય છે અને ARE-gene બેટરીને સક્રિય કરે છે. (B) GSK3 ફોસ્ફોરીલેટ્સ Nrf2 અને આ CUL2-મધ્યસ્થી સર્વવ્યાપકીકરણ અને અનુગામી પ્રોટીઝોમ ડિગ્રેડેશન માટે ?-TrCP દ્વારા Nrf1 ની ઓળખની સુવિધા આપે છે. (C) p62 ને Keap1 સાથે અલગ કરવામાં આવે છે, જે તેના ઓટોફેજિક ડિગ્રેડેશન, Nrf2 ની મુક્તિ અને Nrf2 સિગ્નલિંગમાં વધારો તરફ દોરી જાય છે.
Keap1-સ્વતંત્ર Nrf2 નિયમન
ઉભરતા પુરાવાઓએ Nrf2 નિયમનની એક નવીન પદ્ધતિ જાહેર કરી છે જે Keap1 થી સ્વતંત્ર છે. Nrf6 નું સેરીન-સમૃદ્ધ Neh2 ડોમેન તેના બે હેતુઓ (DSGIS અને DSAPGS) સાથે ?-ટ્રાન્સડ્યુસિન રિપીટ-સમાવતી પ્રોટીન (?-TrCP) [24] સાથે બંધાઈને આ નિયમનમાં નિર્ણાયક ભૂમિકા ભજવે છે. ?-TrCP એ Skp1�Cul1�Rbx1/Roc1 ubiquitin ligase કોમ્પ્લેક્સ માટે સબસ્ટ્રેટ રીસેપ્ટર છે જે Nrf2 ને સર્વવ્યાપકીકરણ અને પ્રોટીસોમલ ડિગ્રેડેશન માટે લક્ષ્ય બનાવે છે. ગ્લાયકોજન સિન્થેઝ કિનેઝ-3 એ Keap1-સ્વતંત્ર Nrf2 સ્થિરીકરણ અને નિયમનમાં સામેલ એક નિર્ણાયક પ્રોટીન છે; તે Nrf2 ને Nrf6 ને ?-TrCP અને ત્યારપછીના પ્રોટીન ડિગ્રેડેશન [2] (ફિગ. 25B) દ્વારા ઓળખવામાં મદદ કરવા માટે Neh1 ડોમેનમાં ફોસ્ફોરીલેટ કરે છે.
અન્ય Nrf2 નિયમનકારો
પુરાવાની બીજી લાઇનએ p62-આશ્રિત Nrf2 સક્રિયકરણનો બિન-પ્રમાણિક માર્ગ જાહેર કર્યો છે જેમાં p62 Keap1 ને ઑટોફેજિક ડિગ્રેડેશન તરફ દોરી જાય છે જે આખરે Nrf2 ના સ્થિરીકરણ અને Nrf2-આશ્રિત જનીનોના ટ્રાન્સએક્ટિવેશન તરફ દોરી જાય છે [26], [27], [28] 29], [1] (ફિગ. XNUMXC).
સંચિત પુરાવા સૂચવે છે કે Nrf2 પ્રવૃત્તિ [30] ના નિયમનમાં ઘણા miRNAs મહત્વપૂર્ણ ભૂમિકા ભજવે છે. સાંગોકોયા એટ અલ. [૩૧] દર્શાવે છે કે miR-31 લિમ્ફોબ્લાસ્ટ K144 સેલ લાઇન, પ્રાથમિક માનવ એરિથ્રોઇડ પૂર્વજ કોષો અને સિકલ-સેલ રોગ રેટિક્યુલોસાઇટ્સમાં Nrf2 પ્રવૃત્તિને સીધી રીતે નિયંત્રિત કરે છે. માનવ સ્તન ઉપકલા કોષોમાં અન્ય એક રસપ્રદ અભ્યાસ દર્શાવે છે કે miR-562 કેપ28-સ્વતંત્ર પદ્ધતિ [2] દ્વારા Nrf1 ને અટકાવે છે. એ જ રીતે, miR-32, miR-153a, miR-27-142p અને miR5 જેવા miRNAs ચેતાકોષીય SH-SY144Y સેલ લાઇન [2] માં Nrf5 અભિવ્યક્તિને નિયંત્રિત કરે છે. સિંઘ વગેરે. [૩૪] દર્શાવે છે કે miR-33 ની એક્ટોપિક અભિવ્યક્તિ 34?-એસ્ટ્રાડિઓલ (E93)-પ્રેરિત ઉંદર મોડેલમાં Nrf2-નિયંત્રિત જનીનની અભિવ્યક્તિને ઘટાડે છે.
અમારી લેબમાંથી તાજેતરની શોધમાં Nrf2 ના અંતર્જાત અવરોધકને ઓળખવામાં આવે છે જે રેટિનોઇક X રીસેપ્ટર આલ્ફા (RXR?) તરીકે ઓળખાય છે. આરએક્સઆર? પરમાણુ રીસેપ્ટર છે, જે Nrf7 (એમિનો-એસિડ અવશેષો 2�209) ના Neh316 ડોમેન સાથે તેના DNA-બંધનકર્તા ડોમેન (DBD) દ્વારા ક્રિયાપ્રતિક્રિયા કરે છે, અને ખાસ કરીને ન્યુક્લિયસમાં Nrf2 પ્રવૃત્તિને અટકાવે છે. તદુપરાંત, અન્ય પરમાણુ રીસેપ્ટર્સ જેમ કે પેરોક્સિસોમ પ્રોલિફેરેટર-એક્ટિવેટેડ રીસેપ્ટર-?, ER?, એસ્ટ્રોજન-સંબંધિત રીસેપ્ટર-?, અને ગ્લુકોકોર્ટિકોઇડ રીસેપ્ટર્સ પણ Nrf2 પ્રવૃત્તિના અંતર્જાત અવરોધક હોવાનું નોંધાયું છે [9], [10].
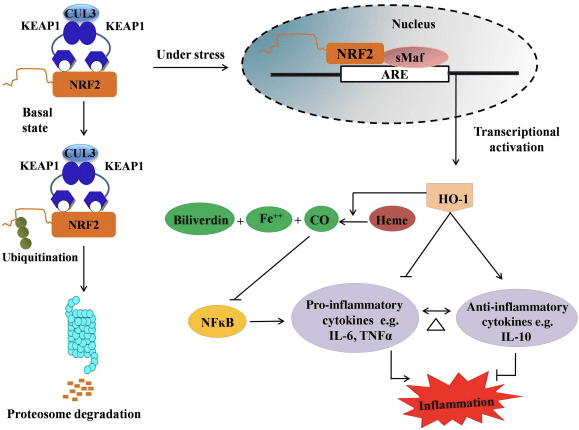
Nrf2/HO-1 એક્સિસની બળતરા વિરોધી ભૂમિકા
HO-1 એ ઇન્ડ્યુસિબલ આઇસોફોર્મ અને દર-મર્યાદિત એન્ઝાઇમ છે જે હેમના કાર્બન મોનોક્સાઇડ (CO) અને ફ્રી આયર્નમાં અને બિલીવર્ડિનને બિલીરૂબિનમાં ઘટાડાનું ઉત્પ્રેરક કરે છે. પ્રો-ઇન્ફ્લેમેટરી ફ્રી હીમનું એન્ઝાઇમેટિક ડિગ્રેડેશન તેમજ CO અને બિલીરૂબિન જેવા બળતરા વિરોધી સંયોજનોનું ઉત્પાદન HO-1 (ફિગ. 2) ની રક્ષણાત્મક અસરોને જાળવવામાં મુખ્ય ભૂમિકા ભજવે છે.
આકૃતિ 2 Nrf2/HO-1 પાથવેનું વિહંગાવલોકન. મૂળભૂત પરિસ્થિતિઓમાં, Nrf2 તેના દબાવનાર Keap1 સાથે જોડાય છે જે સર્વવ્યાપકતા તરફ દોરી જાય છે અને ત્યારબાદ પ્રોટીઝોમ ડિગ્રેડેશન થાય છે. ઓક્સિડેટીવ સ્ટ્રેસ દરમિયાન, ફ્રી Nrf2 ન્યુક્લિયસમાં સ્થાનાંતરિત થાય છે, જ્યાં તે નાના Maf પરિવારના સભ્યો સાથે ડિમરાઇઝ થાય છે અને HO-1 જેવા ARE જનીનો સાથે જોડાય છે. અપરેગ્યુલેટેડ HO-1 હીમને CO, બિલીરૂબિન અને ફ્રી આયર્નમાં ઉત્પ્રેરિત કરે છે. CO એ NF-?B પાથવેના અવરોધક તરીકે કાર્ય કરે છે જે પ્રો-ઈન્ફ્લેમેટરી સાયટોકાઈન્સની ઘટતી અભિવ્યક્તિ તરફ દોરી જાય છે, જ્યારે બિલીરૂબિન એન્ટીઑકિસડન્ટ તરીકે પણ કાર્ય કરે છે. વધુમાં, HO-1 પ્રોઈનફ્લેમેટરી સાયટોકાઈન્સને સીધો અટકાવે છે તેમજ બળતરા વિરોધી સાયટોકાઈન્સને સક્રિય કરે છે, આમ બળતરા પ્રક્રિયાના સંતુલન તરફ દોરી જાય છે.
Nrf2 mRNA અને પ્રોટીન અભિવ્યક્તિ વધારીને HO-1 જનીનને પ્રેરિત કરે છે અને તે ક્લાસિક Nrf2 રેગ્યુલેટેડ જનીનમાંથી એક છે જેનો વ્યાપકપણે અસંખ્ય વિટ્રો અને વિવો અભ્યાસોમાં ઉપયોગ થાય છે. કેટલાક અભ્યાસોએ દર્શાવ્યું છે કે HO-1 અને તેના ચયાપચયમાં Nrf2 દ્વારા મધ્યસ્થી કરવામાં આવતી નોંધપાત્ર બળતરા વિરોધી અસરો છે. HO-1 અભિવ્યક્તિનું એલિવેશન કે જે સક્રિય Nrf2 દ્વારા મધ્યસ્થી કરવામાં આવે છે તે NF?B સિગ્નલિંગના નિષેધ તરફ દોરી જાય છે જેના પરિણામે આંતરડાની મ્યુકોસલ ઈજામાં ઘટાડો થાય છે અને પુરૂષ સ્પ્રેગ-ડોવલી ઉંદર લીવર ટ્રાન્સપ્લાન્ટેશન મોડલ [35] માં ચુસ્ત-જંકશન ડિસફંક્શન થાય છે. Nrf2-આશ્રિત HO-1 અભિવ્યક્તિનું અપગ્ર્યુલેશન H2O12 સાયટોટોક્સિસિટી [2] થી માઉસમાંથી મેળવેલા C2C36 માયોબ્લાસ્ટ્સને સુરક્ષિત કરી શકે છે. Nrf2-આશ્રિત HO-1 એ RAW264.7- અથવા માઉસ પેરીટોનિયલ મેક્રોફેજ-વ્યુત્પન્ન ફોમ સેલ મેક્રોફેજેસમાં લિપોપોલિસેકરાઇડ (LPS) - મધ્યસ્થી બળતરા પ્રતિક્રિયાઓ પર અસર કરે છે. Nrf2 પ્રવૃત્તિ ડિસેન્સિટાઇઝ્ડ ફોમ સેલ મેક્રોફેજ ફેનોટાઇપ અને મેક્રોફેજેસની અમૂલ્ય બળતરાને અટકાવે છે, તે એથરોસ્ક્લેરોસિસ [37] ની પ્રગતિમાં મહત્વપૂર્ણ ભૂમિકા ભજવે છે. Nrf2/HO-1 અક્ષ એલપીએસ પ્રેરિત માઉસ BV2 માઇક્રોગ્લિયલ કોષો અને માઉસ હિપ્પોકેમ્પલ HT22 કોષોને અસર કરે છે, જેની અસર ન્યુરોઇન્ફ્લેમેશન પર થાય છે. માઉસ BV1 માઇક્રોગ્લિયલ કોષોમાં Nrf2 માર્ગ દ્વારા HO-2 અભિવ્યક્તિનું અપગ્ર્યુલેશન જે માઉસ હિપ્પોકેમ્પલ HT22 કોષોના કોષ મૃત્યુને બચાવે છે [38]. વધુમાં, કોબાલ્ટ-આધારિત હાઇબ્રિડ પરમાણુઓ (HYCOs) કે જે Nrf2 ઇન્ડ્યુસરને કાર્બન મોનોક્સાઇડ (CO) ના રીલીઝર સાથે જોડે છે તે Nrf2/HO-1 અભિવ્યક્તિને વધારે છે, CO મુક્ત કરે છે અને વિટ્રોમાં બળતરા વિરોધી પ્રવૃત્તિ કરે છે. HYCOs પણ પેશીઓ HO-1 ને નિયંત્રિત કરે છે અને વિવોમાં વહીવટ પછી રક્તમાં CO પહોંચાડે છે, બળતરા પરિસ્થિતિઓ [39] સામે તેમના સંભવિત ઉપયોગને સમર્થન આપે છે. Nrf2/HO-1 અપરેગ્યુલેશન ટૌરિન ક્લોરામાઇન [40] સાથે સારવાર કરાયેલ મ્યુરિન મેક્રોફેજની ઇફેરોસાયટીક પ્રવૃત્તિમાં વધારો કરીને બળતરા ઘટાડે છે. એકંદરે, ઉપરોક્ત સમજાવેલ પ્રાયોગિક મોડેલો દર્શાવે છે કે Nrf2/HO-1 અક્ષ બળતરા વિરોધી કાર્યમાં મુખ્ય ભૂમિકા ભજવે છે, જે સૂચવે છે કે Nrf2 એ બળતરા-સંબંધિત રોગોમાં ઉપચારાત્મક લક્ષ્ય છે.
વધુમાં, HO-1 ની આડપેદાશો જેમ કે CO, બિલીરૂબિન, ઓક્સિડેટીવ તણાવ અને કોષને નુકસાન દરમિયાન શક્તિશાળી એન્ટીઑકિસડન્ટ તરીકે કામ કરે છે [41], [42]; તે ઓટોઇમ્યુન એન્સેફાલોમીલાઇટિસ અને હેપેટાઇટિસ [43], [44] ને દબાવી દે છે; અને તે iNOS અને NO [45], [46], [47] ની ઉત્પત્તિને અટકાવીને ઉંદર અને ઉંદરોને એન્ડોટોક્સિક આંચકા સામે રક્ષણ આપે છે. વધુમાં, બિલીરૂબિન એન્ડોથેલિયલ એક્ટિવેશન અને ડિસફંક્શનને ઘટાડે છે [48]. રસપ્રદ વાત એ છે કે, બિલીરૂબિન એડહેસન મોલેક્યુલ-1 [49] દ્વારા એન્ડોથેલિયલ લ્યુકોસાઈટ્સના સ્થાનાંતરણને ઘટાડે છે. આ વિશિષ્ટ સંદર્ભો સૂચવે છે કે માત્ર HO-1 બળવાન બળતરા વિરોધી એજન્ટ તરીકે જ નહીં પરંતુ તેના ચયાપચયની ક્રિયાઓ પણ કરે છે.
Nrf2 દ્વારા અવરોધિત બળતરા મધ્યસ્થીઓ અને ઉત્સેચકો
સાયટોકાઇન્સ અને કેમોકાઇન્સ
સાયટોકાઇન્સ એ ઓછા પરમાણુ-વજન પ્રોટીન અને વિવિધ કોષો દ્વારા સ્ત્રાવ થતા પોલિપેપ્ટાઇડ્સ છે; તેઓ કોષની વૃદ્ધિ, ભિન્નતા અને રોગપ્રતિકારક કાર્યને નિયંત્રિત કરે છે અને બળતરા અને ઘા-હીલિંગમાં સામેલ છે. સાયટોકાઇન્સમાં ઇન્ટરલ્યુકિન્સ (ILs), ઇન્ટરફેરોન, ટ્યુમર નેક્રોસિસ ફેક્ટર (TNF), કોલોની-સ્ટિમ્યુલેટિંગ ફેક્ટર, કેમોકાઇન્સ અને વૃદ્ધિ પરિબળોનો સમાવેશ થાય છે. કેટલાક સાયટોકીન્સને બળતરા તરફી મધ્યસ્થીઓ તરીકે ગણવામાં આવે છે જ્યારે અન્યમાં બળતરા વિરોધી કાર્યો હોય છે. ઓક્સિડેટીવ તાણના સંપર્કમાં સાયટોકાઇન્સનું વધુ ઉત્પાદન થાય છે જે લક્ષ્ય કોષોમાં ઓક્સિડેટીવ તણાવનું કારણ બને છે. જ્યારે એનએફ-?બી ઓક્સિડેટીવ સ્ટ્રેસ દ્વારા સક્રિય થાય છે ત્યારે કેટલાક પ્રો-ઇન્ફ્લેમેટરી સાયટોકાઇન્સનું વધુ પડતું ઉત્પાદન થાય છે. વધુમાં, પ્રો-ઈન્ફ્લેમેટરી ઓક્સિડેટીવ તણાવ NF-?B ના વધુ સક્રિયકરણ અને સાયટોકાઈન્સના વધુ ઉત્પાદનનું કારણ બને છે. Nrf2/ARE સિસ્ટમનું સક્રિયકરણ આ ચક્રને વિક્ષેપિત કરવામાં મહત્વપૂર્ણ ભૂમિકા ભજવે છે. કેમોકાઇન્સ એ નાના સાયટોકાઇન્સનું કુટુંબ છે, જેની મુખ્ય ભૂમિકા બળતરા કોશિકાઓના સ્થળાંતરને માર્ગદર્શન આપવાની છે. તેઓ મુખ્યત્વે લ્યુકોસાઈટ્સ, મોનોસાઈટ્સ, ન્યુટ્રોફિલ્સ અને અન્ય ઈફેક્ટર કોષો માટે કીમોએટ્રેક્ટન્ટ તરીકે કાર્ય કરે છે.
એવું નોંધવામાં આવ્યું છે કે Nrf2 નું સક્રિયકરણ IL-6 અને IL-1 સહિત પ્રો-ઇન્ફ્લેમેટરી સાઇટોકીન્સના LPS-પ્રેરિત ટ્રાન્સક્રિપ્શનલ અપગ્ર્યુલેશનને અટકાવે છે? [50]. IL-1? અને IL-6 ઉત્પાદન પણ Nrf2?/? ડેક્સ્ટ્રાન સલ્ફેટ-પ્રેરિત કોલાઇટિસ [51], [52] સાથે ઉંદર. Nrf2 ડાઉનસ્ટ્રીમ IL-17 અને અન્ય બળતરા પરિબળો Th1 અને Th17 ના ઉત્પાદનને અટકાવે છે, અને મલ્ટીપલ સ્ક્લેરોસિસ, ઓટોઇમ્યુન એન્સેફાલીટીસ [53] ના પ્રાયોગિક મોડેલમાં રોગ પ્રક્રિયાને દબાવી દે છે. Nrf2-આશ્રિત એન્ટી-ઓક્સિડન્ટ જનીન HO-1, NQO-1, Gclc, અને Gclm બ્લોક TNF-?, IL-6, મોનોસાઇટ કેમો એટ્રેકન્ટ પ્રોટીન-1 (MCP1), મેક્રોફેજ ઇન્ફ્લેમેટરી પ્રોટીન-2 (MIP2), અને બળતરા મધ્યસ્થી પરંતુ Nrf2-નોકઆઉટ ઉંદરના કિસ્સામાં, બળતરા વિરોધી અસર થતી નથી [54]. LPS સાથે સારવાર કરાયેલ Nrf2-નોકઆઉટ ઉંદરમાંથી પેરીટોનિયલ ન્યુટ્રોફિલ્સમાં વાઇલ્ડ-ટાઇપ (WT) કોષો [6] કરતાં સાયટોકાઇન્સ (TNF-? અને IL-1) અને કેમોકાઇન્સ (MCP2 અને MIP54) નોંધપાત્ર રીતે ઊંચા સ્તરો છે. વિટ્રોમાં, Nrf2 જનીનને માનવ અને સસલાના એઓર્ટિક સ્મૂથ સ્નાયુ કોષોમાં સ્થાનાંતરિત કરવાથી MCP1 [8], [55], અને Nrf2-આશ્રિત HO-1 અભિવ્યક્તિ TNF-?-ઉત્તેજિત NF-?B અને MCP-1 ના સ્ત્રાવને દબાવી દે છે. માનવ નાભિની નસ એન્ડોથેલિયલ કોષોમાં સ્ત્રાવ [56]. આ તારણો સંકેત આપે છે કે, દાહક ઉત્તેજનાના પ્રતિભાવમાં, Nrf2 સિગ્નલિંગનું અપગ્ર્યુલેશન પ્રો-ઈન્ફ્લેમેટરી સાયટોકાઈન્સ અને કેમોકાઈન્સના વધુ પડતા ઉત્પાદનને તેમજ NF-?B ના સક્રિયકરણને મર્યાદિત કરે છે.
કોષ સંલગ્નતા અણુઓ
કોષ સંલગ્નતા અણુઓ (સીએએમ) એ પ્રોટીન છે જે કોષો સાથે અથવા એક્સ્ટ્રા સેલ્યુલર મેટ્રિક્સ સાથે જોડાય છે. કોષની સપાટી પર સ્થિત, તેઓ કોષની ઓળખ, કોષ સક્રિયકરણ, સિગ્નલ ટ્રાન્સડક્શન, પ્રસાર અને ભિન્નતામાં સામેલ છે. CAMs પૈકી, ICAM-1 અને VCAM-1 એ ઇમ્યુનોગ્લોબ્યુલિન સુપરફેમિલીના મહત્વપૂર્ણ સભ્યો છે. ICAM-1 લ્યુકોસાઇટ અને એન્ડોથેલિયલ કોષ પટલમાં ઓછી સાંદ્રતામાં હાજર છે. સાયટોકાઇન ઉત્તેજના પર, સાંદ્રતા નોંધપાત્ર રીતે વધે છે. ICAM-1 ને IL-1 અને TNF દ્વારા પ્રેરિત કરી શકાય છે અને તે વેસ્ક્યુલર એન્ડોથેલિયમ, મેક્રોફેજ અને લિમ્ફોસાઇટ્સ દ્વારા વ્યક્ત થાય છે. તે ઇન્ટિગ્રિન માટે લિગાન્ડ છે, લ્યુકોસાઇટ્સ પર જોવા મળતું રીસેપ્ટર. જ્યારે ICAM-1-ઇંટિગ્રિન બ્રિજ સક્રિય થાય છે, ત્યારે લ્યુકોસાઇટ્સ એન્ડોથેલિયલ કોષો સાથે જોડાય છે અને પછી સબએન્ડોથેલિયલ પેશીઓમાં સ્થળાંતર કરે છે [57]. VCAM-1 લિમ્ફોસાઇટ્સ, મોનોસાઇટ્સ, ઇઓસિનોફિલ્સ અને બેસોફિલ્સને વેસ્ક્યુલર એન્ડોથેલિયમમાં સંલગ્નતામાં મધ્યસ્થી કરે છે અને લ્યુકોસાઇટ ભરતીમાં ફાળો આપે છે, જે આખરે ઓક્સિડેટીવ તણાવને કારણે પેશીઓને નુકસાન તરફ દોરી જાય છે. Nrf2 VCAM-1 [58] ની પ્રમોટર પ્રવૃત્તિને અટકાવે છે. Nrf2-રેગ્યુલેટેડ ડાઉનસ્ટ્રીમ જનીન HO-1, E-selectin અને VCAM-1, એન્ડોથેલિયલ કોષો [59] સાથે સંકળાયેલ સંલગ્ન અણુઓની અભિવ્યક્તિને અસર કરી શકે છે. એનઆરએફ14?/? Nrf1+/+ ઉંદર [1] કરતાં ઉંદર. માનવ મહાધમની એન્ડોથેલિયલ કોષોમાં Nrf2 TNF-?-પ્રેરિત VCAM-2 અભિવ્યક્તિને દબાવી દે છે અને TNF-?-પ્રેરિત મોનોસાયટીક U60 કોષ સંલગ્નતા [2] માં દખલ કરે છે. Nrf1 ની વધુ પડતી અભિવ્યક્તિ માનવ માઇક્રોવેસ્ક્યુલર એન્ડોથેલિયલ કોશિકાઓમાં TNF-?-પ્રેરિત VCAM-937 જનીન અભિવ્યક્તિને પણ અટકાવે છે [8]. કુદરતી રીતે બનતું એન્ટીઑકિસડન્ટ 2-હાઈડ્રોક્સિઆન્થ્રાનિલિક એસિડ (HA), જે બળતરા અથવા ચેપ દરમિયાન kynurenine પાથવે તરીકે ઓળખાતા મેટાબોલિક માર્ગ સાથે વિવોમાં રચાયેલ એલ-ટ્રિપ્ટોફન ચયાપચયમાંથી એક છે, જે HO-1 અભિવ્યક્તિને પ્રેરિત કરે છે અને માનવ નાભિમાં Nrf61 ને ઉત્તેજિત કરે છે. નસ એન્ડોથેલિયલ કોષો (HUVECs). HA દ્વારા પ્રેરિત Nrf3-આશ્રિત HO-1 અભિવ્યક્તિ MCP-2 સ્ત્રાવ, VCAM-2 અભિવ્યક્તિ અને એથરોસ્ક્લેરોસિસ [1] માં વેસ્ક્યુલર ઇજા અને બળતરા સાથે સંકળાયેલ NF-kB સક્રિયકરણને અટકાવે છે. એન્ટિ-પ્રોલિફરેટિવ અને એન્ટિ-ઇન્ફ્લેમેટરી સિન્થેટિક ચૅલકોન ડેરિવેટિવ 1?,1?,56?-ટ્રિસ (મેથોક્સાઇમેથોક્સી) ચેલકોન ICAM-2, પ્રો-ઇન્ફ્લેમેટરી સાઇટોકિન IL-4?, અને TNF-? ટ્રાઇનિટ્રોબેન્ઝીન સલ્ફોનિક એસિડ [6] સાથે સારવાર કરાયેલ ઉંદરમાંથી કોલોનિક પેશીઓમાં અભિવ્યક્તિ. Nrf1 નું અપગ્ર્યુલેશન લાઇકોપીન [1] સાથે સારવાર કરાયેલ માનવ રેટિના રંગદ્રવ્ય ઉપકલા કોષોમાં TNF-?-પ્રેરિત ICAM-62 અભિવ્યક્તિને અટકાવે છે. આ તમામ અભ્યાસો સૂચવે છે કે Nrf2 સોજાની પેશીઓમાં બળતરા કોશિકાઓના સ્થળાંતર અને ઘૂસણખોરીને નિયંત્રિત કરીને બળતરા પ્રક્રિયામાં મુખ્ય ભૂમિકા ભજવે છે.
મેટ્રિક્સ મેટાલોપ્રોટીનેસિસ (MMPs)
MMPs એક્સ્ટ્રા સેલ્યુલર મેટ્રિક્સમાં વ્યાપકપણે હાજર છે અને કોષ પ્રસાર, સ્થળાંતર, ભિન્નતા, ઘા-હીલિંગ, એન્જીયોજેનેસિસ, એપોપ્ટોસિસ અને ટ્યુમર મેટાસ્ટેસિસ જેવી શારીરિક અને રોગવિજ્ઞાન પ્રક્રિયાઓમાં સામેલ છે. એવું નોંધવામાં આવ્યું છે કે Nrf2/HO-1 અક્ષ મેક્રોફેજેસમાં MMP-9 અને માનવ આંતરડાના ઉપકલા કોષોમાં MMP-7 ને અટકાવે છે, અને આ બળતરા આંતરડાના રોગની સારવારમાં ફાયદાકારક છે [62], [64]. યુવી ઇરેડિયેશન-પ્રેરિત ત્વચાને નુકસાન WT ઉંદર કરતાં Nrf2-નોકઆઉટમાં વધુ ગંભીર છે અને MMP-9 સ્તર નોંધપાત્ર રીતે ઊંચું છે, જે દર્શાવે છે કે Nrf2 MMP-9 અભિવ્યક્તિ ઘટાડે છે. તેથી, Nrf2 ને યુવી ઇરેડિયેશન [65] સામે રક્ષણાત્મક માનવામાં આવે છે. અન્ય અભ્યાસમાં એ પણ નોંધવામાં આવ્યું છે કે ટ્યુમર સેલ આક્રમણ અને બળતરામાં MMP-9 નું ડાઉનરેગ્યુલેટેડ ટ્રાન્સક્રિપ્શનલ સક્રિયકરણ NF-kB સિગ્નલિંગ પાથવે [66] ના અવરોધ દ્વારા નિયંત્રિત થાય છે. આઘાતજનક કરોડરજ્જુની ઇજામાં, NF-kB સિગ્નલિંગ પાથવે MMP-9 [67] ના mRNA સ્તરને નિયંત્રિત કરવામાં પણ ભાગ લે છે. તેથી, બળતરામાં MMPsનું નિયમન Nrf2 પાથવે દ્વારા સીધું અથવા Nrf2-પ્રભાવિત NF-?B પાથવે દ્વારા પરોક્ષ રીતે અસર પામે છે.
સાયક્લોક્સીજેનેઝ-2 (COX2) અને ઈન્ડ્યુસિબલ નાઈટ્રિક ઓક્સાઇડ સિન્થેઝ (INOS)
Nrf2-નોકઆઉટ ઉંદર પરના પ્રયોગોની શ્રેણીએ બળતરા અને કોક્સ-2 અને iNOS જેવા પ્રો-ઈન્ફ્લેમેટરી જનીનોના નિયમનમાં તેની નિર્ણાયક ભૂમિકા દર્શાવી છે. પ્રથમ વખત, ખોર એટ અલ. એનઆરએફ2?/? ઉંદર WT Nrf2+/+ ઉંદર સાથે સરખામણી કરે છે, જે દર્શાવે છે કે Nrf2 તેમની પ્રવૃત્તિ [2] દબાવી દે છે. ક્રુસિફેરસ શાકભાજીમાં હાજર જાણીતા Nrf51 એક્ટિવેટર્સ પૈકીના એક, સલ્ફોરાફેન સાથે પ્રીટ્રીટમેન્ટ અંગેના અન્ય અહેવાલમાં, એમઆરએનએ બંને પર TNF-?, IL-2?, COX-1, અને iNOS ની અભિવ્યક્તિને અટકાવવાની તેની બળતરા વિરોધી અસર દર્શાવવામાં આવી છે. અને Nrf2+/+ ઉંદરમાંથી પ્રાથમિક પેરીટોનિયલ મેક્રોફેજમાં પ્રોટીનનું સ્તર Nrf2?/? ઉંદર [2]. એ જ રીતે, LPS-પ્રેરિત બળતરા સાથે Nrf68-નોકઆઉટ ઉંદરનું હિપ્પોકેમ્પસ પણ iNOS, IL-2, અને TNF-? WT ઉંદર [6] કરતાં. તેવી જ રીતે, Nrf69-નોકઆઉટ ઉંદર 2-મિથાઈલ-1-ફિનાઇલ-4-ટેટ્રાહાઇડ્રોપાયરીડિન દ્વારા પ્રેરિત ઓક્સિડેટીવ તણાવ પ્રત્યે અતિસંવેદનશીલ છે તેમજ COX-1,2,3,6, iNOS જેવા બળતરા માર્કર્સના mRNA અને પ્રોટીન સ્તરમાં વધારો દર્શાવે છે. , IL-2, અને TNF-? [6]. વધુમાં, Nrf70?/? મેથિયોનાઇન- અને કોલિન-ઉણપવાળા ખોરાક સાથે પડકારવામાં આવેલા ઉંદરોમાં Cox2 ની mRNA અભિવ્યક્તિ ~ 5-ગણી ઊંચી છે, અને સમાન આહાર પર WT ઉંદરો કરતાં iNOS, Nrf2 [2] ની બળતરા વિરોધી ભૂમિકા સૂચવે છે. તાજેતરમાં, કિમ એટ અલ. એ દર્શાવ્યું કે ફાયટોકેમિકલ એથિલ પાયરુવેટ BV71 કોષોમાં Nrf2 સિગ્નલિંગ દ્વારા iNOS ની અભિવ્યક્તિને ઘટાડીને તેની બળતરા વિરોધી અને વિરોધી ઓક્સિડેટીવ અસરોનો ઉપયોગ કરે છે. તેઓએ બતાવ્યું કે ઇથિલ પાયરુવેટ Nrf2 ના પરમાણુ સ્થાનાંતરણને પ્રેરિત કરે છે, જે આખરે p2 અને p65 વચ્ચેની ક્રિયાપ્રતિક્રિયાને અટકાવે છે, જે iNOS [300] ની અભિવ્યક્તિમાં ઘટાડો તરફ દોરી જાય છે. વધુમાં, કાર્બાઝોલ એનાલોગ LCY-72-CHO Nrf2 ને સક્રિય કરે છે અને તેના પરમાણુ સ્થાનાંતરણનું કારણ બને છે, જે ઉંદરની એઓર્ટિક વેસ્ક્યુલર સ્મૂથ સ્નાયુ કોશિકાઓમાં COX2 અને iNOS અભિવ્યક્તિ [2] ના દમન તરફ દોરી જાય છે.
NLRP2 iIflammasome પ્રવૃતિના નિયમનમાં Nrf3 ની વિરોધાભાસી ભૂમિકા
NLR ફેમિલી, 3 (NLRP3) ઇન્ફ્લેમાસોમ ધરાવતું પાયરીન ડોમેન એક મલ્ટીપ્રોટીન કોમ્પ્લેક્સ છે જે પેથોજેન રેકગ્નિશન રીસેપ્ટર (PRR) તરીકે કાર્ય કરે છે અને માઇક્રોબાયલ, ઓક્સિડેટીવ સ્ટ્રેસ સિગ્નલોની વિશાળ શ્રેણીને ઓળખે છે જેમ કે પેથોજેન-સંબંધિત મોલેક્યુલર પેટર્ન (PAMPs), નુકસાન- સંકળાયેલ મોલેક્યુલર પેટર્ન મોલેક્યુલ્સ (DAMPs) અને ROS [74]. સક્રિય એનએલઆરપી3 ઇન્ફ્લેમસોમ કેસ્પેસ-1 ના ક્લીવેજ અને પ્રો-ઇન્ફ્લેમેટરી સાઇટોકિન ઇન્ટરલ્યુકિન-1 ના સ્ત્રાવમાં મધ્યસ્થી કરે છે? (IL-1?) જે આખરે કોષ મૃત્યુની પ્રક્રિયાને પ્રેરિત કરે છે જેને પાયરોપ્ટોસિસ તરીકે ઓળખવામાં આવે છે જે યજમાનોને પેથોજેન્સની વિશાળ શ્રેણી સામે રક્ષણ આપે છે [75]. જો કે, ઇન્ફ્લેમાસોમનું અવ્યવસ્થિત સક્રિયકરણ પ્રોટીન મિસફોલ્ડિંગ રોગો જેમ કે ટ્રાન્સમિસિબલ સ્પોન્જિફોર્મ એન્સેફાલોપથી, અલ્ઝાઇમર રોગ, પાર્કિન્સન રોગ અને ટાઇપ 2 ડાયાબિટીસ [76], કેન્સર [77], સંધિવા અને એથરોસ્ક્લેરોસિસ [78] સાથે સંકળાયેલું છે.
રોંગ હુ ગ્રૂપના તાજેતરના અવલોકનમાં Nrf2 ના ઇન્ફ્લેમાસોમના નકારાત્મક નિયમન સાથેના જોડાણ પર બહાર આવ્યું છે કે, Nrf2 NQO1 અભિવ્યક્તિને પ્રેરિત કરે છે જે NLRP3 બળતરા સક્રિયકરણ, કેસ્પેસ-1 ક્લીવેજ અને IL-1 ના અવરોધ તરફ દોરી જાય છે? મેક્રોફેજમાં પેઢી. વધુમાં, જાણીતા Nrf2 એક્ટિવેટર, tert-butylhydroquinone (tBHQ) Nrf3-આશ્રિત રીતે ARE ને સક્રિય કરીને NLRP2 ટ્રાન્સક્રિપ્શનને નકારાત્મક રીતે નિયંત્રિત કરે છે [79]. ઉપરોક્ત અવલોકન ઉપરાંત, તે જ જૂથ એ પણ બહાર આવ્યું છે કે, ડાઇમેથાઇલ ફ્યુમરેટ (DMF) Nrf2 સિગ્નલિંગ પાથવેને સક્રિય કરીને DSS-પ્રેરિત કોલાઇટિસને અટકાવે છે જે Nrf2 પરમાણુ ટ્રાન્સલોકેશન અને NLRP3 ઇન્ફ્લેમાસોમ એસેમ્બલી [80] ના અવરોધમાં સામેલ છે.
કુદરતી અને કૃત્રિમ સંયોજનોનો ઉપયોગ કરીને પ્રયોગોની શ્રેણીએ NLRP2 બળતરા સક્રિયકરણ પર Nrf3 ની અવરોધક અસર પણ જાહેર કરી છે. દાખલા તરીકે, લ્યુપસ નેફ્રાઇટિસ ઉંદરમાં એપિગાલોકેટેચીન-3-ગેલેટ (EGCG) ની સારવારમાં રેનલ NLRP3 ઇન્ફ્લેમસોમ સક્રિયકરણમાં ઘટાડો જોવા મળ્યો છે જે Nrf2 સિગ્નલિંગ પાથવે [81] દ્વારા મધ્યસ્થી થાય છે. તેવી જ રીતે, સિટ્રાલ (3,7-ડાઇમિથાઇલ-2,6-ઓક્ટેડિનલ), ચાઇનીઝ હર્બલ મેડિસિન લિટ્સિયા ક્યુબેબામાં એક મુખ્ય સક્રિય સંયોજન છે, જે એક્સિલરેટેડ અને સિવિયર લ્યુપસ નેફ્રાઇટિસ (ASLN) મોડલમાં Nrf3 એન્ટીઑકિસડન્ટ સિગ્નલિંગ પાથવે દ્વારા NLRP2 બળતરા સક્રિયકરણને અટકાવે છે. [82]. એ જ રીતે, બાયોકેનિન Nrf2 પાથવેને સક્રિય કરીને અને NLRP3 નર BALB/c ઉંદર [83] માં બળતરાના સક્રિયકરણને અટકાવીને LPS/GalN-પ્રેરિત યકૃતની ઇજા સામે રક્ષણ આપે છે. વધુમાં, મેંગીફેરીનને ડોઝ-આશ્રિત રીતે Nrf2 અને HO-1 ની અભિવ્યક્તિને નિયંત્રિત કરવા માટે પણ દર્શાવવામાં આવ્યું હતું અને LPS/D-GalN-પ્રેરિત યકૃત NLRP3, ASC, caspase-1, IL-1 ને અવરોધે છે? અને TNF-? અભિવ્યક્તિ [84].
Nrf3 દ્વારા NLRP2 ના નકારાત્મક નિયમન છતાં, તે NLRP3 અને AIM2 બળતરા કાર્યને પણ સક્રિય કરે છે. હૈતાઓ વેન અને સહકર્મીઓએ શોધ્યું કે, Nrf2 ?/? માઉસ મેક્રોફેજે NLRP3 અને AIM2 ઈન્ફ્લેમસોમનું ખામીયુક્ત સક્રિયકરણ દર્શાવ્યું છે પરંતુ NLRC4 ઈન્ફ્લેમસોમ [85] નથી. રસપ્રદ રીતે, આ અવલોકન બળતરા સંબંધિત રોગોના સંદર્ભમાં Nrf2 ના અજાણ્યા કાર્યોનું નિરૂપણ કરે છે; આથી ઉપચારાત્મક લક્ષ્ય તરીકે વિચારતા પહેલા Nrf2 બળતરાના કાર્યને સક્રિય કરે છે તે પદ્ધતિને જાહેર કરવા માટે વધુ અભ્યાસ કરવો ખૂબ જ મહત્વપૂર્ણ છે.
Nrf2 દ્વારા પ્રો-ઈન્ફ્લેમેટરી સાયટોકાઈન ટ્રાન્સક્રિપ્શનનું દમન
માઉસ મેક્રોફેજેસમાં ક્રોમેટિન ઇમ્યુનોપ્રિસિપિટેશન (ChIP)-seq અને ChIP-qPCR પર આધારિત ખૂબ જ તાજેતરની તપાસમાં બહાર આવ્યું છે કે Nrf2 IL-6 અને IL-1 જેવા પ્રો-ઇન્ફ્લેમેટરી સાઇટોકાઇન્સના પ્રમોટર પ્રદેશો સાથે જોડાય છે? અને RNA Pol II ભરતીને અટકાવે છે. પરિણામે, RNA Pol II IL-6 અને IL-1 ના ટ્રાન્સક્રિપ્શનલ સક્રિયકરણની પ્રક્રિયા કરવામાં અસમર્થ છે? જે આખરે જનીન અભિવ્યક્તિના અવરોધ તરફ દોરી જાય છે. પ્રથમ વખત, મસાયુકી યામામોટોના જૂથે નવલકથા પદ્ધતિનો ખુલાસો કર્યો જેના દ્વારા Nrf2 માત્ર તેના ડાઉનસ્ટ્રીમ જનીનોને AREs દ્વારા જ નહીં પરંતુ RNA Pol II [50] ની ભરતીને અટકાવીને ARE સાથે અથવા તેના વગર ચોક્કસ જનીનોના ટ્રાન્સક્રિપ્શનલ સક્રિયકરણને પણ દબાવી દે છે.
Nrf2 અને NF-?B પાથવેઝ વચ્ચે ક્રોસસ્ટૉક
NF-?B એ DNA ટ્રાન્સક્રિપ્શન માટે જવાબદાર પ્રોટીન સંકુલ છે જે લગભગ તમામ પ્રકારના પ્રાણી કોષોમાં જોવા મળે છે અને વિવિધ પ્રક્રિયાઓ જેમ કે બળતરા, એપોપ્ટોસીસ, રોગપ્રતિકારક પ્રતિક્રિયા, કોષની વૃદ્ધિ અને વિકાસમાં સામેલ છે. p65, NF-?B કુટુંબનું એક Rel પ્રોટીન, એક ટ્રાન્સએક્ટિવેશન ડોમેન ધરાવે છે જ્યારે p50 નથી કરતું અને ટ્રાન્સક્રિપ્શનને સક્રિય કરવા માટે Rel પ્રોટીન સાથે હેટરોડાઈમરાઈઝેશનની જરૂર છે. ઓક્સિડેટીવ તણાવ દરમિયાન, I?B કિનેઝ (IKK) સક્રિય થાય છે અને I?B ના ફોસ્ફોરાયલેશનનું કારણ બને છે, પરિણામે NF-?B ના પ્રકાશન અને પરમાણુ સ્થાનાંતરણ થાય છે. NF-?B IL-6, TNF-?, iNOS, IL-1, અને અંતઃકોશિક સંલગ્નતા COX-2 જેવા બળતરા તરફી મધ્યસ્થીઓના ટ્રાન્સક્રિપ્શનનું કારણ બને છે.
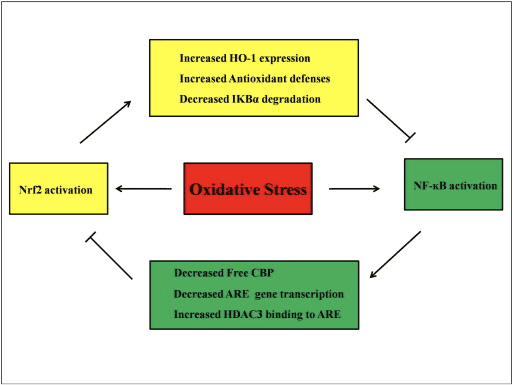
NF-?B નું અસામાન્ય નિયમન સંધિવા, અસ્થમા, આંતરડાના દાહક રોગ અને હેલિકોબેક્ટર પાયલોરી ચેપ-પ્રેરિત ગેસ્ટ્રાઇટિસ [86] સાથે જોડાયેલું છે. હાલમાં એવું માનવામાં આવે છે કે NF-kB પ્રવૃત્તિ Keapl/Nrf2/ARE સિગ્નલિંગ પાથવેને મુખ્યત્વે ત્રણ પાસાઓમાં પ્રભાવિત કરે છે: પ્રથમ, Keap1 IKK ને અધોગતિ કરે છે? સર્વવ્યાપકતા દ્વારા, આમ NF-?B [87] ની પ્રવૃત્તિને અવરોધે છે. બીજું, દાહક પ્રક્રિયા સાયક્લોપેન્ટેનોન પ્રોસ્ટાગ્લાન્ડિન 2d-PGJ15 માંથી મેળવેલા COX2 જેવા બળતરા મધ્યસ્થીઓને પ્રેરિત કરે છે, જે એક મજબૂત ઇલેક્ટ્રોફાઇલ છે જે Keap1 સાથે પ્રતિક્રિયા આપે છે અને Nrf2 ને સક્રિય કરે છે, આમ NF-kB (58) પ્રવૃત્તિના એક સાથે અવરોધ સાથે જીન ટ્રાન્સક્રિપ્શન શરૂ કરે છે. ફિગ. 88 A, B). ત્રીજું, NF-?B સ્પર્ધાત્મક Nrf3 ટ્રાન્સક્રિપ્શનલ કો-એક્ટિવેટર CBP [2], [89] (ફિગ. 90 C, D) સાથે જોડાઈ શકે છે.
આકૃતિ 3 Nrf2 અને NF-?B માર્ગો વચ્ચેનો ક્રોસસ્ટાલ્ક. (A) Keap1 IKK ને CUL3-મધ્યસ્થી સર્વવ્યાપકીકરણ અને પ્રોટીઝોમ ડિગ્રેડેશન તરફ દોરી જાય છે જે આખરે NF-?B ફોસ્ફોરાયલેશનના નિષેધ તરફ દોરી જાય છે અને આ પદ્ધતિ Keap2 સાથે Nrf1 અને IKK ના સ્પર્ધાત્મક બંધન તરીકે પણ કામ કરે છે. (B) ઓક્સિડેટીવ તણાવ IKK ને સક્રિય કરે છે જે NF-?B ને ફોસ્ફોરીલેટ કરે છે, જે તેના ન્યુક્લિયસમાં સ્થાનાંતરણ તરફ દોરી જાય છે અને કોક્સ-2 જેવા પ્રોઇનફ્લેમેટરી સાયટોકાઇન્સને સક્રિય કરે છે. COX-2 નું ટર્મિનલ ઉત્પાદન 15d-PGJ2 તરીકે ઓળખાય છે તે Nrf2 ના પ્રેરક તરીકે કાર્ય કરે છે જે આખરે ઓક્સિડેટીવ તણાવના દમન તરફ દોરી જાય છે. (C) Nrf2 એ આરઇ-સંચાલિત જનીન અભિવ્યક્તિ શરૂ કરવા માટે નાના Maf અને અન્ય ટ્રાન્સક્રિપ્શનલ મશીનરી સાથે તેના ટ્રાન્સક્રિપ્શનલ કોફેક્ટર CBP સાથે જોડાય છે. (D) જ્યારે NF-?B સ્પર્ધાત્મક રીતે CBP સાથે જોડાય છે, ત્યારે તે CBP ને Nrf2 સાથે બંધનને અટકાવે છે, જે Nrf2 ટ્રાન્સએક્ટિવેશનના નિષેધ તરફ દોરી જાય છે.
એવું માનવામાં આવે છે કે Nrf2 અને NF-?B સિગ્નલિંગ પાથવે ડાઉનસ્ટ્રીમ લક્ષ્ય પ્રોટીનના ટ્રાન્સક્રિપ્શન અથવા કાર્યને નિયંત્રિત કરવા માટે ક્રિયાપ્રતિક્રિયા કરે છે. આ ધારણાના સમર્થનમાં ઘણા ઉદાહરણો દર્શાવે છે કે Nrf2 અને NF-?B માર્ગો (ફિગ. 4). LPS ના પ્રતિભાવમાં, Nrf2 નોકડાઉન નોંધપાત્ર રીતે NF-?B ટ્રાન્સક્રિપ્શનલ પ્રવૃત્તિ અને NF-?B-આશ્રિત જીન ટ્રાન્સક્રિપ્શનમાં વધારો કરે છે, જે દર્શાવે છે કે Nrf2 NF-?B પ્રવૃત્તિ [60], [91] ને અવરોધે છે. વધુમાં, Nrf2-આશ્રિત ડાઉનસ્ટ્રીમ HO-1 ની વધેલી અભિવ્યક્તિ NF-?B પ્રવૃત્તિને અટકાવે છે. જ્યારે પ્રોસ્ટેટ કેન્સરના કોષો સંક્ષિપ્તમાં ?-ટોકોફેરિલ સસીનેટના સંપર્કમાં આવે છે, જે વિટામિન ઇનું વ્યુત્પન્ન છે, HO-1 અભિવ્યક્તિ અપરેગ્યુલેટ થાય છે. HO-1 ના અંતિમ ઉત્પાદનો NF-?B [92] ના પરમાણુ સ્થાનાંતરણને અટકાવે છે. આ વિવો અભ્યાસ સૂચવે છે કે Nrf2 નકારાત્મક રીતે NF-kB સિગ્નલિંગ પાથવેનું નિયમન કરે છે. LPS NF-?B DNA બંધનકર્તા પ્રવૃત્તિને ઉત્તેજિત કરે છે અને NF-?B ના p65 સબ્યુનિટનું સ્તર Nrf2?/? ના ફેફસાંમાંથી અણુ અર્કમાં નોંધપાત્ર રીતે વધારે છે. WT ઉંદર કરતાં, NF-?B સક્રિયકરણમાં Nrf2 ની નકારાત્મક ભૂમિકા સૂચવે છે. વધુમાં, Nrf2?/? LPS અને TNF- સાથે માઉસ એમ્બ્રીયો ફાઈબ્રોબ્લાસ્ટની સારવાર કરવામાં આવે છે? IKK સક્રિયકરણ અને I?B-? અધોગતિ [60]. અને Nrf2?/? ઉંદર WT ઉંદર [93] સાથે સરખામણી કરે છે. Nrf2 માં પ્રિસ્ટેન-પ્રેરિત લ્યુપસ નેફ્રાઇટિસ?/? સલ્ફોરાફેન સાથે સહ-સારવાર કરાયેલ ઉંદરને ગંભીર રેનલ નુકસાન અને પેથોલોજીકલ ફેરફારો તેમજ WT ની સરખામણીમાં એલિવેટેડ iNOS અભિવ્યક્તિ અને NF-?B સક્રિયકરણ હોય છે, જે સૂચવે છે કે Nrf2 NF-?B સિગ્નલિંગ પાથવેને અટકાવીને અને ROS [94 સાફ કરીને લ્યુપસ નેફ્રાઇટિસને સુધારે છે. ]. NF-?B પ્રવૃત્તિ પણ ત્યારે થાય છે જ્યારે કોષોને LPS અને TNF-? સાથે Nrf2 પ્રેરક સાથે સારવાર આપવામાં આવે છે. ઉદાહરણ તરીકે, કૃત્રિમ ચેલકોન ડેરિવેટિવ TNF-?-પ્રેરિત NF-?B સક્રિયકરણને પ્રત્યક્ષ અને પરોક્ષ રીતે અને અંશતઃ માનવ આંતરડાના ઉપકલા HT-1 કોષોમાં HO-29 અભિવ્યક્તિના ઇન્ડક્શન દ્વારા અટકાવે છે [62]. જ્યારે F344 ઉંદરોને 3H-1,2-dithiole-3-thione (D3T) [95] સાથે સારવાર આપવામાં આવે છે ત્યારે NF-?B ટ્રાન્સલોકેશન અને DNA-બંધનકર્તા પ્રવૃત્તિનું દમન તેમજ હિપેટોસાઇટ્સમાં iNOS અભિવ્યક્તિનું દમન જોવા મળે છે. સલ્ફોરાફેન અને LPS સાથે સહ-સારવાર પછી, iNOS, COX-2, અને TNF-ની LPS-પ્રેરિત અભિવ્યક્તિ? કાચા 264.7 માં મેક્રોફેજનું નિયમન ઓછું થાય છે, સૂચવે છે કે સલ્ફોરાફેન એનએફ-?બી ડીએનએ બંધનકર્તા [96] ના અવરોધ દ્વારા બળતરા વિરોધી પ્રવૃત્તિ ધરાવે છે. Nrf2 અને NF-?B માર્ગો વચ્ચેની લિંકને સમજાવવા માટે ઘણા પ્રાયોગિક અભ્યાસો કરવામાં આવ્યા હોવા છતાં, વિરોધાભાસી પરિણામો રહે છે. Nrf2 અને NF-kB [97] વચ્ચે હકારાત્મક અને નકારાત્મક બંને નિયમનો નોંધવામાં આવ્યા છે. સામાન્ય રીતે, કેમોપ્રિવેન્ટિવ ઇલેક્ટ્રોફાઇલ્સ 3H-1,2-ડિથિઓલ-3-થિઓન, સલ્ફોરાફેન અને ટ્રાઇટરપેનોઇડ CDDO-Me NF-kB અને તેના ડાઉનરેગ્યુલેટેડ જનીનો [2], [98], [99] ને અટકાવીને Nrf100 ને સક્રિય કરે છે. તેનાથી વિપરિત, ROS, LPS, ફ્લો શીયર સ્ટ્રેસ, ઓક્સિડાઇઝ્ડ LDL અને સિગારેટનો ધુમાડો જેવા કેટલાક એજન્ટો અથવા સ્થિતિઓ Nrf2 અને NF-kB પ્રવૃત્તિ [97] બંનેમાં વધારો કરવા માટે દર્શાવવામાં આવ્યા છે. વધુમાં, વિવોના અભ્યાસમાં જાણવા મળ્યું છે કે Nrf2?/? ઉંદર અને NF-?B બંધનકર્તા પ્રવૃત્તિ Nrf2 માં ઓછી છે?/? Nrf2+/+ ઉંદર [101] કરતાં. જો કે, એડીનોવાયરલ વેક્ટર Nrf2 સાથે સારવાર કરાયેલ માનવ એઓર્ટિક એન્ડોથેલિયલ કોષો NF-?B [8] ની પ્રવૃત્તિને અસર કર્યા વિના NF-?B ડાઉનસ્ટ્રીમ જનીનોને અટકાવે છે.
આકૃતિ 4 Nrf2 અને NF-?B નો નિયમનકારી લૂપ. Nrf2 પાથવે I?B- ના અધોગતિને અટકાવીને NF-?B સક્રિયકરણને અટકાવે છે. અને HO-1 અભિવ્યક્તિ અને એન્ટીઑકિસડન્ટ સંરક્ષણમાં વધારો જે આરઓએસ અને ડિટોક્સિફાઇંગ રસાયણોને તટસ્થ કરે છે. પરિણામે, ROS-સંબંધિત NF-?B સક્રિયકરણ દબાવવામાં આવે છે. તેવી જ રીતે, NF-?B- મધ્યસ્થી ટ્રાંસ્ક્રિપ્શન� ઘટાડીને Nrf2 સક્રિયકરણ ઘટાડે છેછેCBP માટે Nrf2 સાથે સ્પર્ધા કરીને જીન ટ્રાન્સક્રિપ્શન અને મફત CREB બંધનકર્તા પ્રોટીન. વધુમાં, NF-?B ARE પ્રદેશમાં હિસ્ટોન ડીસીટીલેઝ (HDAC3) ની ભરતીમાં વધારો કરે છે અને તેથી Nrf2 ટ્રાન્સક્રિપ્શનલ સક્રિયકરણ અટકાવવામાં આવે છે.
Nrf2 સિગ્નલિંગ પાથવેનું સક્રિયકરણ માનવ શરીરમાં કોશિકાઓની એન્ટીઑકિસડન્ટ ક્ષમતાને વધારીને પ્રતિક્રિયાશીલ ઑક્સિડન્ટ્સના ડિટોક્સિફિકેશનમાં સામેલ ઉત્સેચકો અને જનીનોની અભિવ્યક્તિમાં મુખ્ય ભૂમિકા ભજવે છે. જ્યારે આજે ઘણા સંશોધન અભ્યાસો ઉપલબ્ધ છે, Nrf2 સક્રિયકરણમાં નિયમનકારી પદ્ધતિઓ સંપૂર્ણપણે સમજી શકાતી નથી. બળતરાની સારવારમાં Nrf2 સિગ્નલિંગ પાથવેની સંભવિત ભૂમિકા પણ મળી આવી છે. ડૉ. એલેક્સ જિમેનેઝ ડીસી, CCST ઇનસાઇટ
બળતરા રોગોમાં Nrf2 ની ભૂમિકા
વિવોના અભ્યાસોએ દર્શાવ્યું છે કે Nrf2 વિવિધ સિસ્ટમોને અસર કરતા બળતરા રોગોમાં મહત્વની ભૂમિકા ભજવે છે; આમાં ગેસ્ટ્રાઇટિસ, કોલાઇટિસ, આર્થરાઇટિસ, ન્યુમોનિયા, લીવર ડેમેજ, કાર્ડિયોવેસ્ક્યુલર ડિસીઝ, ન્યુરોડિજનરેટિવ ડિસીઝ અને મગજને નુકસાનનો સમાવેશ થાય છે. આ અભ્યાસોમાં, Nrf2?/? ડબલ્યુટી પ્રાણીઓ કરતાં પ્રાણીઓએ બળતરા અને પેશીઓને નુકસાનના વધુ ગંભીર લક્ષણો દર્શાવ્યા હતા. તેથી, એવું માનવામાં આવે છે કે Nrf2 સિગ્નલિંગ પાથવે બળતરા રોગોમાં રક્ષણાત્મક અસર ધરાવે છે. પોર્સિન સ્વાદુપિંડના ઇલાસ્ટેઝનું ઇન્ટ્રા-ટ્રાચેલ ઇન્સ્ટોલેશન ક્રોનિક અવરોધક પલ્મોનરી રોગ, ખાસ કરીને એમ્ફિસીમાને પ્રેરિત કરે છે. Nrf2-ઉણપવાળા ઉંદર એમ્ફિસીમા માટે અત્યંત સંવેદનશીલ હોય છે, અને HO-1, PrxI, અને એન્ટિપ્રોટીઝ જનીન SLPI ની અભિવ્યક્તિમાં ઘટાડો મૂર્ધન્ય મેક્રોફેજમાં થાય છે. Nrf2 ને ફેફસાની ઇજા [102] સામે મેક્રોફેજ મધ્યસ્થી સંરક્ષણ પ્રણાલીમાં મુખ્ય નિયમનકાર તરીકે ગણવામાં આવે છે. 2 મહિના સુધી તમાકુના ધુમાડાના સંપર્કથી પ્રેરિત એમ્ફિસીમા સાથે Nrf6-ઉણપવાળા ઉંદર બ્રોન્કોઆલ્વિઓલર સોજામાં વધારો દર્શાવે છે, એલ્વિઓલીમાં ઓક્સિડેટીવ સ્ટ્રેસ માર્કર્સની અપરેગ્યુલેટેડ અભિવ્યક્તિ અને મૂર્ધન્ય સેપ્ટલ સેલ એપોપ્ટોસીસમાં વધારો દર્શાવે છે, જે સૂચવે છે કે Nrf2 એમ્ફિસીમા સામે કૃત્રિમ રીતે વધેલા એમ્ફિસીમા સામે અસર કરે છે. જનીનો [102], [103]. Nrf2 વિક્ષેપ સાથે, ઓવલબ્યુમિન કોમ્પ્લેક્સનો ઉપયોગ કરીને એલર્જન-મધ્યસ્થી વાયુમાર્ગમાં બળતરા અને અસ્થમા વાયુમાર્ગની બળતરા, વાયુમાર્ગની હાયપર-રિએક્ટિવિટી, ગોબ્લેટ કોશિકાઓના હાયપરપ્લાસિયા અને બ્રોન્કોઆલ્વીઓલર લેવેજ અને સ્પ્લેનોસાઇટ્સમાં Th2 નું ઉચ્ચ સ્તર દર્શાવે છે, જ્યારે Nrf2-મધ્યસ્થી સિગ્નલ લિમિટેડ એરવેમાં વધારો દર્શાવે છે. , મ્યુકસ હાઇપરસેક્રેશન, અને એરવે હાઇપર-રિએક્ટિવિટી તેમજ ઘણા એન્ટીઑકિસડન્ટ જનીનોને પ્રેરિત કરે છે જે અસ્થમાના વિકાસને અટકાવે છે [104]. પ્લ્યુરલ કેવિટીમાં કેરેજીનન ઈન્જેક્શન પ્યુરીસીને પ્રેરિત કરે છે, અને Nrf15 બળતરા કોશિકાઓમાં 2d-PGJ2 સંચય માઉસ પેરીટોનિયલ મેક્રોફેજ સુધી મર્યાદિત છે. બળતરાના પ્રારંભિક તબક્કા દરમિયાન, 15d-PGJ2 Nrf2 ને સક્રિય કરે છે અને HO-1 અને PrxI ના ઇન્ડક્શન દ્વારા બળતરા પ્રક્રિયાને નિયંત્રિત કરે છે. એક અભ્યાસમાં એવું પણ સૂચવવામાં આવ્યું છે કે COX-2 15d-PGJ2 [105] ના ઉત્પાદન દ્વારા પ્રારંભિક તબક્કામાં બળતરા વિરોધી અસર ધરાવે છે. 1 અઠવાડિયા માટે 1% ડેક્સ્ટ્રાન સલ્ફેટ સોડિયમનું મૌખિક વહીવટ હિસ્ટોલોજીકલ ફેરફારો સાથે સંકળાયેલ કોલાઇટિસને પ્રેરિત કરે છે જેમાં ક્રિપ્ટ્સનું શોર્ટનિંગ અને કોલોન પેશીઓમાં બળતરા કોશિકાઓની ઘૂસણખોરીનો સમાવેશ થાય છે. કોલાઇટિસમાં આંતરડાની અખંડિતતાને સુરક્ષિત કરવા માટે, Nrf2 પ્રો-ઇન્ફ્લેમેટરી સાઇટોકીન્સનું નિયમન કરીને અને તબક્કા II ડિટોક્સિફાઇંગ એન્ઝાઇમ્સને પ્રેરિત કરીને મહત્વપૂર્ણ ભૂમિકા ભજવી શકે છે [51]. LPS-પ્રેરિત પલ્મોનરી સેપ્સિસના Nrf2-નોકઆઉટ માઉસ મોડેલમાં, NF-?B પ્રવૃત્તિ કોક્સ-2, IL-113, IL-6, અને TNF જેવા બળતરા સાઇટોકીન્સના પ્રભાવને નિયંત્રિત કરે છે? જે બળતરા શરૂ કરવા અને પ્રોત્સાહન આપવા માટે જરૂરી છે [60]. Nrf2 આ દાહક પરિબળોને નિયંત્રિત કરીને દાહક નુકસાન ઘટાડે છે. તીવ્ર બળતરાના આ મોડેલોમાં, Nrf2 સિગ્નલિંગ પાથવે દ્વારા એન્ટીઑકિસડન્ટ એન્ઝાઇમ્સ, પ્રો-ઇન્ફ્લેમેટરી સાયટોકાઇન્સ અને મધ્યસ્થીઓનું વધતું નિયમન WT પ્રાણીઓમાં બળતરાની ઇજાને ઘટાડે છે. રસપ્રદ વાત એ છે કે, આ Nrf2-નોકઆઉટ ઉંદરમાં પણ નોંધવામાં આવ્યું છે જેમાં ડબ્લ્યુટી ઉંદરની તુલનામાં લક્ષણો નોંધપાત્ર રીતે વધારે છે.
Nrf2-આશ્રિત બળતરા વિરોધી દવાઓ પર સંશોધન
સારાંશમાં, અમે પ્રયોગોની ચર્ચા કરી છે જે દર્શાવે છે કે Nrf2 સિગ્નલ પાથવે બળતરાના ઘણા ક્ષેત્રોમાં નિયમનકારી ભૂમિકા ભજવે છે, તેથી Nrf2-આશ્રિત બળતરા વિરોધી એજન્ટો બળતરા રોગોની સારવાર માટે મહત્વપૂર્ણ છે.
છોડ એ સંયોજનોના અસાધારણ રીતે સમૃદ્ધ સ્ત્રોત છે જે Nrf2 ટ્રાન્સક્રિપ્શન પરિબળને સક્રિય કરે છે, જે સાયટોપ્રોટેક્ટીવ જનીનોના નિયમન તરફ દોરી જાય છે. તાજેતરમાં, વિવિધ બળતરા વિરોધી એજન્ટોની અસરોની તપાસ કરવા માટે ઘણા અભ્યાસો હાથ ધરવામાં આવ્યા હતા, મોટાભાગે છોડના મૂળના. ઉદાહરણ તરીકે, કર્ક્યુમિન હળદરનું સક્રિય ઘટક છે અને તે આદુમાં પણ ઓછી માત્રામાં જોવા મળે છે; આઇસોથિયોસાયનેટ્સ, ખાસ કરીને ફેનીલિસોથિયોસાયનેટ્સ બ્રોકોલી, સેલરી અને અન્ય શાકભાજીમાંથી છે; અને એન્થોકયાનિન બેરી અને દ્રાક્ષમાંથી મળે છે [124]. અભ્યાસોએ દર્શાવ્યું છે કે આ તમામ એજન્ટો માત્ર સારા એન્ટીઑકિસડન્ટો નથી પણ Nrf2 ઇન્ડક્શન [125], [126] દ્વારા બળવાન બળતરા વિરોધી અસરો પણ ધરાવે છે. તેથી, છોડના અર્કમાંથી નવા એન્ટિ-ઇન્ફ્લેમેટરી Nrf2 એક્ટિવેટર્સના વિકાસે તબીબી સંશોધનમાં ખૂબ જ રસ આકર્ષ્યો છે.
તાજેતરના વર્ષોમાં, આ સંયોજનોની ક્રિયાઓની પુષ્ટિ કરવા માટે ઘણા પ્રાણી પ્રયોગો હાથ ધરવામાં આવ્યા છે. આર્ટેસુનેટનો ઉપયોગ મુખ્યત્વે ગંભીર મેલેરિયા, સેરેબ્રલ મેલેરિયા અને સંધિવાયુક્ત સ્વયંપ્રતિરક્ષા રોગો માટે થાય છે; તે સેપ્ટિક ફેફસાની ઇજામાં પણ અસરકારક છે. આર્ટેસુનેટ Nrf2 અને HO-1 અભિવ્યક્તિને સક્રિય કરે છે, અને બાદમાં બળતરાને રોકવા માટે પેશીઓમાં પ્રો-ઇન્ફ્લેમેટરી સાયટોકાઇન્સ અને લ્યુકોસાઇટ્સના પ્રવાહને ઘટાડે છે [127]. આઇસોવિટેક્સિન, ઓરીઝા સટીવા ચોખાના હલમાંથી કાઢવામાં આવે છે, એવું માનવામાં આવે છે કે તે બળતરા વિરોધી અને એન્ટીઑકિસડન્ટ ગુણધર્મો ધરાવે છે; તે Nrf2/HO-1 પાથવેને સક્રિય કરીને અને MAPK અને NF-?B [128] ને અટકાવીને LPS-પ્રેરિત તીવ્ર ફેફસાની ઈજા સામે રક્ષણાત્મક ભૂમિકા ભજવે છે. ફિમાસર્ટન, રેનિન-એન્જિયોટેન્સિન સિસ્ટમ પર કામ કરતું નવું લોકપ્રિય એન્જીયોટેન્સિન II રીસેપ્ટર બ્લોકર, બ્લડ પ્રેશર ઘટાડે છે; શસ્ત્રક્રિયા-પ્રેરિત એકપક્ષીય મૂત્રમાર્ગ અવરોધ સાથે ઉંદરની સારવાર માટે ફિમાસર્ટનનો ઉપયોગ Nrf2 અને એન્ટીઑકિસડન્ટ પાથવેને અપરેગ્યુલેટ કરીને અને RAS અને MAPKs [129] ને અટકાવીને ઓક્સિડેટીવ તણાવ, બળતરા અને ફાઇબ્રોસિસ ઘટાડે છે. સપાનોન દક્ષિણપૂર્વ એશિયામાં વ્યાપકપણે વિતરિત થાય છે, જ્યાં તેનો ઉપયોગ એન્ટી-ઈન્ફલ્યુએન્ઝા, એન્ટિ-એલર્જિક અને ન્યુરોપ્રોટેક્ટીવ દવા તરીકે થાય છે; તે Nrf2 ને સક્રિય કરે છે અને NF-?B ને અટકાવે છે અને તેથી Nrf2- અને/અથવા NF-?B- સંબંધિત રોગો [130] ની સારવારમાં ફાયદાકારક હોઈ શકે છે. Bixin orellana ના બીજમાંથી કાઢવામાં આવેલ Bixin નો ઉપયોગ મેક્સિકો અને દક્ષિણ અમેરિકામાં ચેપી અને બળતરા રોગો માટે થાય છે; તે વેન્ટિલેશન-પ્રેરિત ફેફસાની ઇજાને દૂર કરવા અને સામાન્ય ફેફસાના મોર્ફોલોજી [2]ને પુનઃસ્થાપિત કરવા Nrf131-આશ્રિત રીતે બળતરા મધ્યસ્થીઓ, મૂર્ધન્ય કેશિલરી લિકેજ અને ઓક્સિડેટીવ નુકસાન ઘટાડે છે. અન્ય છોડના સંયોજનો, જેમ કે એપીગાલોકેટેચીન ગેલેટ, સલ્ફોરાફેન, રેઝવેરાટ્રોલ, લાઇકોપીન અને ગ્રીન ટી અર્ક, Nrf2 સિગ્નલિંગ પાથવે [132], [133], [134] દ્વારા બળતરા રોગો પર ઉપચારાત્મક અસર કરે છે. તાજેતરમાં અન્ય ફાયટોકેમિકલ, એરિઓડિક્ટિઓલ, જે સાઇટ્રસ ફળોમાં હાજર છે, તે Nrf2 ને નિયંત્રિત કરીને, NF-?B ને અટકાવીને સિસ્પ્લેટિન-પ્રેરિત કિડનીની ઇજા અને સેપ્સિસ-પ્રેરિત તીવ્ર ફેફસાની ઇજા પર બળતરા વિરોધી અને એન્ટીઑકિસડન્ટ અસરો હોવાનું નોંધવામાં આવ્યું છે. મેક્રોફેજમાં સાયટોકાઈન્સની અભિવ્યક્તિ [135], [136]. જો કે, અસંખ્ય ફાયટોકેમિકલ્સ વિવિધ માનવ રોગોની રોકથામ અને સારવાર માટે મહાન વચન દર્શાવે છે, અને કેટલાક પહેલેથી જ ક્લિનિકલ ટ્રાયલ તબક્કામાં પ્રવેશી ચૂક્યા છે (કોષ્ટક 2).
આ પ્લાન્ટ સંયોજનો Nrf2 સિગ્નલિંગ પાથવેને મુખ્યત્વે ઇલેક્ટ્રોફિલિક સામગ્રીના સ્વરૂપમાં સક્રિય કરે છે જે Keap1 ના સિસ્ટીન અવશેષોને સંશોધિત કરે છે, જે ARE સાથે મુક્ત પરમાણુ Nrf2 બંધન તરફ દોરી જાય છે, જેના પરિણામે સંબંધિત જનીનનું ટ્રાન્સક્રિપ્શન સક્રિય થાય છે.
સલ્ફોરાફેન અને કેન્સર, મૃત્યુદર, વૃદ્ધત્વ, મગજ અને વર્તન, હૃદય રોગ અને વધુ પર તેની અસરો
Isothiocyanates એ કેટલાક સૌથી મહત્વપૂર્ણ છોડ સંયોજનો છે જે તમે તમારા આહારમાં મેળવી શકો છો. આ વિડિયોમાં હું તેમના માટે અત્યાર સુધીનો સૌથી વ્યાપક કેસ બનાવું છું. ટૂંકા ધ્યાન ગાળો? નીચેના ટાઈમ પોઈન્ટ્સમાંથી કોઈ એક પર ક્લિક કરીને તમારા મનપસંદ વિષય પર જાઓ. નીચે સંપૂર્ણ સમયરેખા.
મુખ્ય વિભાગો:
00:01:14 - કેન્સર અને મૃત્યુદર
00:19:04 - વૃદ્ધત્વ
00:26:30 - મગજ અને વર્તન
00:38:06 - અંતિમ રીકેપ
00:40:27 - માત્રા
સંપૂર્ણ સમયરેખા:
00:00:34 - સલ્ફોરાફેનનો પરિચય, વિડીયોનું મુખ્ય ધ્યાન.
00:01:14 - ક્રુસિફેરસ શાકભાજીનો વપરાશ અને સર્વ-કારણ મૃત્યુદરમાં ઘટાડો.
00:44:24 - શાકભાજીમાંથી સક્રિય માયરોસિનેઝ સાથે જોડવામાં આવે ત્યારે પૂરક વધુ સારી રીતે કામ કરે છે.
00:44:56 - રાંધવાની તકનીકો અને ક્રુસિફેરસ શાકભાજી.
00:46:06 - ગોઇટ્રોજન તરીકે આઇસોથિયોસાયનેટ્સ.
નિષ્કર્ષ
હાલમાં, ઘણા સંશોધનોએ બળતરામાં Nrf2/Keap1/ARE સિગ્નલિંગ પાથવેની ભૂમિકા પર ધ્યાન કેન્દ્રિત કર્યું છે. Nrf2 દ્વારા અપરેગ્યુલેટ કરાયેલા ઉત્સેચકોમાં, HO-1 પ્રતિનિધિ તણાવ પ્રતિભાવ ઉત્સેચકોમાંનું એક છે. HO-1 અગ્રણી બળતરા વિરોધી અને એન્ટીઑકિસડન્ટ ગુણધર્મો ધરાવે છે. સામાન્ય રીતે, Nrf2 સિગ્નલિંગ પાથવે સાયટોકાઇન્સ, કેમોકિન મુક્ત કરનારા પરિબળો, MMPs અને અન્ય બળતરા મધ્યસ્થીઓ COX-2 અને iNOS ઉત્પાદનને પણ નકારાત્મક રીતે નિયંત્રિત કરે છે, જે સંબંધિત NF-kB અને MAPK માર્ગો અને અન્ય નેટવર્કને સીધી કે પરોક્ષ રીતે અસર કરે છે જે બળતરાને નિયંત્રિત કરે છે. એવું સૂચવવામાં આવે છે કે Nrf2 અને NF-?B સિગ્નલિંગ પાથવે ડાઉનસ્ટ્રીમ લક્ષ્ય પ્રોટીનના ટ્રાન્સક્રિપ્શન અથવા કાર્યને નિયંત્રિત કરવા માટે ક્રિયાપ્રતિક્રિયા કરે છે. Nrf2 દ્વારા NF-?B- મધ્યસ્થી ટ્રાન્સક્રિપ્શનલ પ્રવૃત્તિનું દમન અથવા નિષ્ક્રિયકરણ કદાચ બળતરાના પ્રારંભિક તબક્કામાં થાય છે, કારણ કે NF-?B બળતરા તરફી મધ્યસ્થીઓની શ્રેણીના ડી નોવો સંશ્લેષણને નિયંત્રિત કરે છે. જો કે, સંશોધનમાં હજુ પણ કેટલીક મર્યાદાઓ છે જેમ કે Nrf2 અને અન્ય સિગ્નલિંગ માર્ગો જેમ કે JAK/STAT વચ્ચે જોડાણો છે કે કેમ, બળતરામાં કુદરતી વનસ્પતિ સ્ત્રોતોમાંથી મેળવેલા વર્તમાન Nrf2 એક્ટિવેટર્સનું મહત્વ અને જૈવિક પ્રવૃત્તિને કેવી રીતે સુધારવી. અને આ સંયોજનોના લક્ષ્યાંકને વધારે છે. આને વધુ પ્રાયોગિક માન્યતાની જરૂર છે.
વધુમાં, Nrf2 સિગ્નલિંગ પાથવે > 600 જનીનોને નિયંત્રિત કરી શકે છે [163], જેમાંથી > 200 એન્કોડ સાયટોપ્રોટેક્ટીવ પ્રોટીન [164] કે જે બળતરા, કેન્સર, ન્યુરોડિજનરેટિવ રોગો અને અન્ય મોટા રોગો [165] સાથે પણ સંકળાયેલા છે. વધતા જતા પુરાવા સૂચવે છે કે, Nrf2 સિગ્નલિંગ પાથવે ઘણા કેન્સરોમાં અંકુશમુક્ત છે, જેના પરિણામે Nrf2 આધારિત જનીન બેટરી અસ્પષ્ટ અભિવ્યક્તિમાં પરિણમે છે. વધુમાં, બળતરા ઓક્સિડેટીવ તણાવ સંબંધિત રોગોમાં ખાસ કરીને કેન્સરમાં મુખ્ય ભૂમિકા ભજવે છે. બળતરાનો સામનો કરવા માટે ઘણા Nrf2 એક્ટિવેટર્સનો ઉપયોગ Nrf2 ડાઉનસ્ટ્રીમ જનીનોની અસ્પષ્ટ અભિવ્યક્તિમાં પરિણમી શકે છે જે ઓન્કોજેનેસિસ અને કીમો અને/અથવા રેડિયો થેરાપી સામે પ્રતિકાર પ્રેરિત કરે છે. તેથી, તેની પ્લિયોટ્રોપિક અસરોને ઘટાડવા માટે Nrf2 ના અત્યંત વિશિષ્ટ એક્ટિવેટર્સ વિકસાવવામાં આવી શકે છે. Nrf2 ના કેટલાક સક્રિયકર્તાઓએ ઓક્સિડેટીવ તણાવ સંબંધિત રોગોમાં બળતરા વિરોધી કાર્યોમાં નોંધપાત્ર સુધારો દર્શાવ્યો છે. એફડીએ દ્વારા મંજૂર કરાયેલ અને મલ્ટીપલ સ્ક્લેરોસિસ (એમએસ) જેવા દાહક રોગની સારવાર માટે વ્યાપકપણે ઉપયોગમાં લેવાતા Nrf2 એક્ટિવેટરનું શ્રેષ્ઠ ઉદાહરણ છે ડાયમિથાઈલ ફ્યુમરેટ. મોટી સંખ્યામાં દર્દીઓ [152] માં મલ્ટીપલ સ્ક્લેરોસિસના રિલેપ્સિંગ સ્વરૂપોની સારવાર માટે ટેકફિડેરા (બાયોજેન દ્વારા ડાયમિથાઈલ ફ્યુમરેટનું નોંધાયેલ નામ) અસરકારક રીતે ઉપયોગમાં લેવાય છે. જો કે, બળતરા રોગોની સારવાર માટે Nrf2 એક્ટિવેટર્સનો ઉપયોગ કરવાની અસરકારકતાને Nrf2 ની હાનિકારક અસરોને ટાળવા માટે વધુ માન્યતાની જરૂર છે. તેથી, Nrf2 દ્વારા મધ્યસ્થી કરવામાં આવતી બળતરા વિરોધી પ્રવૃત્તિ માટે ઉપચારનો વિકાસ નોંધપાત્ર ક્લિનિકલ અસર કરી શકે છે. સમગ્ર વિશ્વમાં Nrf2 સિગ્નલિંગ પાથવેના ચાલુ અભ્યાસો બળતરાના લક્ષણોને નિયંત્રિત કરવા અને કેન્સર તેમજ ન્યુરોડિજનરેટિવ અને અન્ય મુખ્ય રોગોને રોકવા અને સારવાર માટે ઉચ્ચ-લક્ષિત ઉપચારાત્મક એજન્ટો વિકસાવવા માટે સમર્પિત છે.
નિષ્કર્ષમાં, Nrf2 માનવ શરીરમાં ઓક્સિડેટીવ સ્ટ્રેસના સ્તરને સમજે છે અને અંતે એન્ટીઑકિસડન્ટ અને ડિટોક્સિફાઇંગ એન્ઝાઇમ્સ અને જનીનોના નિયમનને પ્રોત્સાહન આપવામાં મદદ કરે છે. કારણ કે ઓક્સિડેટીવ સ્ટ્રેસના વધતા સ્તરને કારણે થતી ક્રોનિક સોજા ન્યુરોડિજનરેટિવ રોગો સાથે સંકળાયેલી છે, Nrf2 આવશ્યક ભૂમિકા ભજવી શકે છે અલ્ઝાઈમર રોગ જેવી સ્વાસ્થ્ય સમસ્યાઓની સારવારમાં, અન્યો વચ્ચે. અમારી માહિતીનો અવકાશ શિરોપ્રેક્ટિક અને કરોડરજ્જુના સ્વાસ્થ્ય સમસ્યાઓ સુધી મર્યાદિત છે. વિષયની ચર્ચા કરવા માટે, કૃપા કરીને ડૉ. જીમેનેઝને પૂછો અથવા અમારો સંપર્ક કરો915-850-0900 .
ઘૂંટણની પીડા એ જાણીતું લક્ષણ છે જે ઘૂંટણની વિવિધ ઇજાઓ અને/અથવા પરિસ્થિતિઓને કારણે થઈ શકે છે, જેમાં�રમતો ઇજાઓ. ઘૂંટણ એ માનવ શરીરના સૌથી જટિલ સાંધાઓમાંનું એક છે કારણ કે તે ચાર હાડકાં, ચાર અસ્થિબંધન, વિવિધ રજ્જૂ, બે મેનિસ્કી અને કોમલાસ્થિના આંતરછેદથી બનેલું છે. અમેરિકન એકેડેમી ઓફ ફેમિલી ફિઝિશિયન્સ અનુસાર, ઘૂંટણની પીડાના સૌથી સામાન્ય કારણોમાં પેટેલર સબલક્સેશન, પેટેલર ટેન્ડિનિટિસ અથવા જમ્પર્સ ઘૂંટણ અને ઓસ્ગુડ-સ્લેટર રોગનો સમાવેશ થાય છે. જોકે ઘૂંટણનો દુખાવો 60 વર્ષથી વધુ ઉંમરના લોકોમાં થવાની સંભાવના છે, પરંતુ ઘૂંટણનો દુખાવો બાળકો અને કિશોરોમાં પણ થઈ શકે છે. ઘૂંટણના દુખાવાની સારવાર RICE પદ્ધતિઓને અનુસરીને ઘરે કરી શકાય છે, જો કે, ઘૂંટણની ગંભીર ઇજાઓને તાત્કાલિક તબીબી સહાયની જરૂર પડી શકે છે, જેમાં શિરોપ્રેક્ટિક સંભાળનો સમાવેશ થાય છે. �
ન્યુરોડિજનરેટિવ રોગો, જેમ કે અલ્ઝાઈમર રોગ અને પાર્કિન્સન રોગ, વિશ્વભરમાં લાખો વ્યક્તિઓને અસર કરે છે. ઘણા ન્યુરોડિજનરેટિવ રોગોના લક્ષણોની સારવાર માટે વિવિધ સારવાર વિકલ્પો ઉપલબ્ધ છે, જોકે પરિણામો ઘણીવાર મર્યાદિત હોય છે. સંશોધન અભ્યાસમાં જાણવા મળ્યું છે કે આંતરિક અને બાહ્ય બંને પરિબળોને કારણે ઓક્સિડેટીવ તણાવ ન્યુરોડિજનરેટિવ રોગોના વિકાસનું કારણ બની શકે છે. આ ટ્રાન્સક્રિપ્શન પરિબળ, Nrf2, ઓક્સિડેટીવ તાણ સામે મુખ્ય સંરક્ષણ પદ્ધતિ તરીકે કાર્ય કરવા માટે નક્કી કરવામાં આવ્યું છે. નીચેના લેખનો હેતુ ની અસરો બતાવવાનો છે ન્યુરોડીજનરેટિવ રોગો પર Nrf2.
ટ્રાન્સક્રિપ્શન ફેક્ટર NRF2 દ્વારા પ્રોટીઓસ્ટેસિસનું મોડ્યુલેશન
ન્યુરોડિજનરેટિવ રોગો ચોક્કસ પ્રોટીન એકત્રીકરણ સાથે સંકળાયેલા છે, જે ઇજાગ્રસ્ત મગજ અને પ્રોટીઓસ્ટેસિસના નુકશાન વચ્ચે ઘનિષ્ઠ જોડાણ સૂચવે છે. પ્રોટીઓસ્ટેસીસ એ તમામ પ્રક્રિયાઓનો ઉલ્લેખ કરે છે જેના દ્વારા કોષો પ્રોટીઓમની વિપુલતા અને ફોલ્ડિંગને નિયંત્રિત કરે છે અને વિશાળ નેટવર્કને આભારી છે જે સિગ્નલિંગ પાથવેઝ, જનીન અભિવ્યક્તિ અને પ્રોટીન ડિગ્રેડેશન સિસ્ટમ્સના નિયમનને એકીકૃત કરે છે. આ સમીક્ષા ટ્રાન્સક્રિપ્શન ફેક્ટર NRF2 (પરમાણુ પરિબળ (erythroid-derived 2)-like 2) દ્વારા કરવામાં આવેલ પ્રોટીઓસ્ટેસિસના ટ્રાન્સક્રિપ્શનલ મોડ્યુલેશન વિશેના સૌથી સુસંગત તારણોનો સારાંશ આપવાનો પ્રયાસ કરે છે. NRF2 ને શાસ્ત્રીય રીતે એન્ટીઑકિસડન્ટ સેલ પ્રતિભાવના મુખ્ય નિયમનકાર તરીકે ગણવામાં આવે છે, જો કે તે હાલમાં પ્રોટીઓસ્ટેસિસ જાળવવા ટ્રાન્સડક્શન મશીનરીના મુખ્ય ઘટક તરીકે ઉભરી રહ્યું છે. જેમ જેમ આપણે ચર્ચા કરીશું તેમ, NRF2 ની કલ્પના એક કેન્દ્ર તરીકે કરી શકાય છે જે સંકલિત અને ટકાઉ ટ્રાન્સક્રિપ્શનલ પ્રતિભાવ બનાવવા માટે ખોટી ફોલ્ડ કરેલ પ્રોટીન સંચયમાંથી મેળવેલા કટોકટી સંકેતોનું સંકલન કરે છે. એન્ડોપ્લાઝમિક રેટિક્યુલમ ફિઝિયોલોજી, પ્રોટીઝોમ અને ઓટોફેજીની જાળવણીમાં સંકળાયેલા જનીનોના નિયંત્રણ સાથે સંબંધિત NRF2 ના કાર્યો દ્વારા આ પ્રાપ્ત થાય છે.
ન્યુક્લિયર ફેક્ટર (erythroid-derived 2)-like 2 (NRF2) એ મૂળભૂત-લ્યુસીન-ઝિપર પ્રોટીન છે જેને આજકાલ સેલ્યુલર હોમિયોસ્ટેસિસના મુખ્ય નિયમનકાર તરીકે ગણવામાં આવે છે. તે 250 થી વધુ જનીનોની મૂળભૂત અને તાણ-ઇન્દ્યુસિબલ અભિવ્યક્તિને નિયંત્રિત કરે છે જે એન્ટિઓક્સિડન્ટ રિસ્પોન્સ એલિમેન્ટ (ARE) [1], [2], [3], [4], [5] તરીકે ઓળખાતા સીઆઈએસ-એક્ટિંગ એન્હાન્સર સમાન રીતે વહેંચે છે. આ જનીનો તબક્કા I, II અને III બિનઝેરીકરણ પ્રતિક્રિયાઓ, ગ્લુટાથિઓન અને પેરોક્સિરેડોક્સિન/થિયોરેડોક્સિન ચયાપચય, પેન્ટોઝ ફોસ્ફેટ માર્ગ દ્વારા NADPH ઉત્પાદન અને મેલિક એન્ઝાઇમ, ફેટી એસિડ ઓક્સિડેશન, આયર્ન ચયાપચય અને પ્રોટીઓસ્ટેસિસ [6] માં ભાગ લે છે. આ વ્યાપક સાયટોપ્રોટેક્ટીવ કાર્યોને જોતાં, શક્ય છે કે NRF2 માં એક જ ફાર્માકોલોજિકલ હિટ ઓક્સિડેટીવ, બળતરા અને પ્રોટીટોક્સિક તણાવ સહિત ક્રોનિક રોગોના મુખ્ય ગુનેગારોની અસરને ઘટાડી શકે. એન્ટીઑકિસડન્ટ સંરક્ષણના મોડ્યુલેશન અને બળતરાના ઉકેલમાં NRF2 ની ભૂમિકા અસંખ્ય અભ્યાસોમાં સંબોધવામાં આવી છે ([7] માં સમીક્ષા). અહીં, અમે પ્રોટીઓસ્ટેસિસમાં તેની ભૂમિકા પર ધ્યાન કેન્દ્રિત કરીશું, એટલે કે, પ્રોટીન સંશ્લેષણ, ફોલ્ડિંગ, હેરફેર અને ડિગ્રેડેશનના હોમિયોસ્ટેટિક નિયંત્રણ. ન્યુરોડિજનરેટિવ રોગોના સંદર્ભમાં ઉદાહરણો આપવામાં આવશે.
પ્રોટીઓસ્ટેસિસનું નુકસાન ન્યુરોડિજનરેટિવ રોગોમાં NRF2 પ્રવૃત્તિને પ્રભાવિત કરે છે
ન્યુરોડિજનરેટિવ રોગોની સામાન્ય ઓળખ એ અમુક પ્રોટીનનું અવ્યવસ્થિત એકત્રીકરણ છે. આમ, પાર્કિન્સન રોગ (PD), ?-amyloid (A?) તકતીઓ અને અલ્ઝાઈમર રોગ (AD) માં હંટીંગટીન (Htt) માં હાઇપર-ફોસ્ફોરીલેટેડ TAU ન્યુરોફિબ્રિલરી ટેંગલ્સમાં ?-synuclein (?-SYN) ના મિસફોલ્ડ પ્રોટીન એગ્રીગેટ્સ જોવા મળે છે. હંટીંગ્ટન ડિસીઝ (HD), સુપરઓક્સાઇડ ડિસમ્યુટેઝ 1 (SOD1) અને TAR DNA બાઈન્ડિંગ પ્રોટીન 43 (TDP-43), એમિઓટ્રોફિક લેટરલ સ્ક્લેરોસિસ (ALS), સ્પોન્જિફોર્મ એન્સેફાલોપથીમાં પ્રિઓન પ્રોટીન (PrP), વગેરે. પ્રોટીન એગ્રીગેટ્સ અનેક પર અસર કરી શકે છે. સેલ્યુલર માર્ગો, જે બદલામાં NRF2 સ્તર અને પ્રવૃત્તિને અસર કરી શકે છે.
નિયમનના વિવિધ સ્તરો NRF2 પ્રવૃત્તિને ચુસ્તપણે નિયંત્રિત કરે છે
શારીરિક પરિસ્થિતિઓ હેઠળ, કોષો તેના ઝડપી ટર્નઓવરને કારણે નીચા NRF2 પ્રોટીન સ્તરો દર્શાવે છે. વિવિધ ઉત્તેજનાના પ્રતિભાવમાં, NRF2 પ્રોટીન સંચિત થાય છે, ન્યુક્લિયસમાં પ્રવેશ કરે છે અને ARE ધરાવતા જનીનોનું ટ્રાન્સક્રિપ્શન વધારે છે. તેથી, NRF2 પ્રોટીન સ્તરોનું સંચાલન એ એક મુખ્ય મુદ્દો છે જે હકારાત્મક અને નકારાત્મક ઇનપુટ સિગ્નલોને એકીકૃત કરવા જોઈએ. જેમ જેમ આપણે આગળ ચર્ચા કરીશું, NRF2 એ ઝડપી અને કાર્યક્ષમ પ્રતિભાવ આપવા માટે વિવિધ ઓવરલેપિંગ મિકેનિઝમ્સ દ્વારા સક્રિય કરવામાં આવે છે પરંતુ બીજી તરફ NRF2 ને અટકાવી શકાય છે, કદાચ બીજા તબક્કામાં, તેના પ્રતિભાવને બંધ કરવા માટે.
ક્લાસિક દૃષ્ટિકોણથી, NRF2 ના સક્રિયકરણને ઓક્સિડન્ટ અથવા ઇલેક્ટ્રોફિલિક સંયોજનો માટે સેલ્યુલર પ્રતિભાવના પરિણામ તરીકે ગણવામાં આવે છે. આ સંદર્ભમાં, ubiquitin E3 ligase એડેપ્ટર Kelch-like ECH-સંબંધિત પ્રોટીન 1 (KEAP1) નિર્ણાયક ભૂમિકા ભજવે છે. મોલેક્યુલર વિગતો વિભાગ 4.1 માં વધુ સંબોધવામાં આવશે. સંક્ષિપ્તમાં, KEAP1 એ NRF2 સર્વવ્યાપકતા અને પ્રોટીસોમલ ડિગ્રેડેશન તરફ દોરી રહેલા ગંભીર સિસ્ટીન અવશેષોને કારણે રેડોક્સ સેન્સર તરીકે કાર્ય કરે છે. આ ક્લાસિક મોડ્યુલેશન ઉપરાંત, NRF2 સિગ્નલિંગ ઈવેન્ટ્સ દ્વારા ગહનપણે નિયંત્રિત થાય છે. ખરેખર, NRF2 ને ફોસ્ફોરીલેટ અને નિયમન કરવા માટે વિવિધ કિનાસ દર્શાવવામાં આવ્યા છે. દાખલા તરીકે, NRF2 ને મિટોજન એક્ટિવેટેડ પ્રોટીન કિનાસિસ (MAPKs) દ્વારા ફોસ્ફોરીલેટેડ કરી શકાય છે, જોકે NRF2 પ્રવૃત્તિમાં તેનું યોગદાન અસ્પષ્ટ રહે છે [8], [9], [10], [11]. PKA કિનાઝ તેમજ કેટલાક PKC આઇસોઝાઇમ્સ [12], CK2 [13] અથવા Fyn [14] ફોસ્ફોરીલેટ NRF2 તેની સ્થિરતામાં ફેરફાર કરે છે. અમારા જૂથના પાછલા કાર્યએ અહેવાલ આપ્યો કે ગ્લાયકોજેન સિન્થેઝ કિન્સે -3? (GSK-3?) NRF2 ને પરમાણુ બાકાત અને પ્રોટીસોમલ ડિગ્રેડેશન [15], [25], [26], [27], [28], [29], [30] દ્વારા અટકાવે છે. પરમાણુ વિગતોની ચર્ચા વિભાગ 4.1 માં કરવામાં આવશે. વધુમાં, NRF2 અન્ય પ્રકારના નિયમન માટે સબમિટ કરવામાં આવે છે. દા.ત. [૧૮].
NRF2 રેગ્યુલેટરી મિકેનિઝમ્સ પર પ્રોટીન એગ્રીગેટ્સની અસર
આ વિભાગમાં આપણે ધ્યાન કેન્દ્રિત કરીશું કે કેવી રીતે ખોટી ફોલ્ડ કરેલ પ્રોટીનનું સંચય NRF2 પ્રવૃત્તિને અસર કરી શકે છે જે ઉપર દર્શાવેલ કેટલાક માર્ગો ઉદાહરણરૂપ ઉદાહરણ તરીકે પ્રદાન કરે છે. સૌપ્રથમ, આપણે એ ધ્યાનમાં લેવાની જરૂર છે કે પ્રોટીન સંચય ઓક્સિડેટીવ નુકસાન સાથે સખત રીતે જોડાયેલું છે. ખરેખર, મિટોકોન્ડ્રિયા અને અન્ય સ્ત્રોતો [19] માંથી મિસફોલ્ડ પ્રોટીન સંચય અને એકત્રીકરણ પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓ (ROS) ના અસામાન્ય ઉત્પાદનને પ્રેરિત કરે છે. ઉપર જણાવ્યા મુજબ, ROS KEAP1 ના રેડોક્સ-સંવેદનશીલ સિસ્ટીનને સંશોધિત કરશે જે NRF2 ના પ્રકાશન, સ્થિરીકરણ અને પરમાણુ સ્થાનિકીકરણ તરફ દોરી જશે.
પ્રોટીનોપેથી વિશે, NRF2 ને અસર કરી શકે તેવી અવ્યવસ્થિત સિગ્નલિંગ ઘટનાઓનું ઉદાહરણ GSK-3 ના હાયપરએક્ટિવેશન દ્વારા પ્રદાન કરવામાં આવ્યું છે? AD માં. GSK-3?, જેને TAU kinase તરીકે પણ ઓળખવામાં આવે છે, તે આ માઇક્રોટ્યુબ્યુલ-સંબંધિત પ્રોટીનના ફોસ્ફોરાયલેશનમાં ભાગ લે છે, જેના પરિણામે તેનું એકત્રીકરણ, ન્યુરોફિબ્રિલરી ટેંગલ્સની રચના અને એક્સોનલ ટ્રાન્સપોર્ટમાં વિક્ષેપ થાય છે ([20] માં સમીક્ષા કરવામાં આવી છે). બીજી બાજુ, GSK-3? ઉપર જણાવ્યા મુજબ NRF2 સ્તર અને પ્રવૃત્તિને નાટકીય રીતે ઘટાડે છે. વ્યાપકપણે સ્વીકૃત ન હોવા છતાં, એમીલોઇડ કાસ્કેડ પ્રસ્તાવ કરે છે કે ઝેરી A? ઓલિગોમર્સ GSK-3 વધારશે? TAU હાયપર-ફોસ્ફોરાયલેશન અને ન્યુરોન ડેથ [21], [22] સાથે મળીને પ્રવૃત્તિ. એ કેવી રીતે સમજાવવા માટે વિવિધ મોડેલો છે? GSK3-ની તરફેણ કરે છે? પ્રવૃત્તિ. દાખલા તરીકે, એ? ઇન્સ્યુલિન રીસેપ્ટર સાથે જોડાય છે અને PI3K અને AKT સિગ્નલિંગ પાથવેને અટકાવે છે, જે GSK-3 જાળવવા માટે નિર્ણાયક છે? તેના N-ટર્મિનલ Ser9 અવશેષો [23] પર ફોસ્ફોરાયલેશન દ્વારા નિષ્ક્રિય. બીજી બાજુ, બાહ્યકોષીય એ? Frizzled રીસેપ્ટર્સ સાથે ક્રિયાપ્રતિક્રિયા કરે છે, WNT સિગ્નલિંગ [24] ને અવરોધે છે અને ફરીથી સક્રિય GSK-3 ના પ્રકાશનમાં પરિણમે છે. સારાંશમાં, એ? સંચય GSK-3? ના અસામાન્ય હાયપરએક્ટિવેશન તરફ દોરી જાય છે, આમ યોગ્ય NRF2 પ્રતિભાવને બગાડે છે.
નીચેના વિભાગમાં ચર્ચા કર્યા મુજબ, ખોટી ફોલ્ડ કરેલ પ્રોટીન PERK અને MAPK ના સક્રિયકરણ તરફ દોરી જાય છે, જે બદલામાં NRF2 [31], [8], [9], [10], [11] ને નિયંત્રિત કરે છે. વધુમાં, અસંયમિત CBP/p300 પ્રવૃત્તિ અનેક પ્રોટીનોપેથીઓમાં નોંધવામાં આવી છે [૩૨] અને એડી મગજમાં ડીએનએ મેથિલેશનમાં વૈશ્વિક ઘટાડો પણ દર્શાવવામાં આવ્યો હતો [૩૩], તેથી NRF32 નિયમનમાં આ તારણોની સુસંગતતા શોધવા માટે આધાર પૂરો પાડે છે.
અમે અને અન્ય લોકોએ PD અને AD દર્દીઓના નેક્રોપ્સીમાં NRF2 પ્રોટીન સ્તરમાં વધારો અને તેના કેટલાક લક્ષ્યો, જેમ કે હેમ ઓક્સિજેનેઝ 1 (HMOX1), NADPH ક્વિનોન ઓક્સિડેઝ 1 (NQO1), p62, વગેરે, ઇમ્યુનોબ્લોટ અને બંને દ્વારા જોવા મળ્યું છે. ઇમ્યુનોહિસ્ટોકેમિસ્ટ્રી [34], [35], [36], [37], [38], [39] દ્વારા. આ રોગોમાં NRF2 ના અપ-રેગ્યુલેશનને હોમિયોસ્ટેટિક મૂલ્યોને પુનઃપ્રાપ્ત કરવાના રોગગ્રસ્ત મગજના અસફળ પ્રયાસ તરીકે અર્થઘટન કરવામાં આવે છે. જો કે, અન્ય અભ્યાસ દર્શાવે છે કે NRF2 મુખ્યત્વે AD હિપ્પોકેમ્પલ ચેતાકોષોના સાયટોપ્લાઝમમાં સ્થાનીકૃત છે, જે મગજમાં NRF2 ટ્રાન્સક્રિપ્શનલ પ્રવૃત્તિમાં ઘટાડો સૂચવે છે [40]. તે કલ્પનાશીલ છે કે આ અવલોકનોની અસમાનતા ન્યુરોડિજનરેશનના પ્રગતિશીલ તબક્કાઓ સાથે NRF2 ને નિયંત્રિત કરતા પરિબળોમાં ફેરફાર સાથે સંબંધિત છે.
ત્રણ મુખ્ય પ્રણાલીઓ પ્રોટીઓસ્ટેસિસમાં ફાળો આપે છે, જેમ કે અનફોલ્ડ પ્રોટીન રિસ્પોન્સ (UPR), યુબીક્વિટીન પ્રોટીઝોમ સિસ્ટમ (યુપીએસ) અને ઓટોફેજી. આગળ, અમે પ્રોટીન ડેરિવેટિવ મશીનરી સાથે પ્રોટીન એગ્રીગેટ્સ દ્વારા સક્રિય કરાયેલા કટોકટી સંકેતોને જોડતા હબ તરીકે NRF2 ની કલ્પના કરવાના પુરાવા રજૂ કરીએ છીએ.
NRF2 અનફોલ્ડ પ્રોટીન રિસ્પોન્સ (UPR) માં ભાગ લે છે
યુપીઆરના પ્રતિભાવમાં NRF2 સક્રિયકરણ
ER માં ઓક્સિડેટીવ પ્રોટીન ફોલ્ડિંગ સંખ્યાબંધ વિશિષ્ટ માર્ગો દ્વારા ચલાવવામાં આવે છે, જેમાં સૌથી વધુ સંરક્ષિત પ્રોટીન ડાયસલ્ફાઇડ-આઇસોમેરેઝ (PDI) અને સલ્ફાઇડ્રિલ ઓક્સિડેઝ એન્ડોપ્લાઝમિક ઓક્સિડોરેડક્ટીન 1 (ERO1? અને ERO1? સસ્તન પ્રાણીઓમાં)નો સમાવેશ થાય છે. સંક્ષિપ્તમાં, PDI તેના પોતાના સિસ્ટીન એમિનોએસિડ્સના ઘટાડા અને ઓક્સિડેશનને કારણે, પ્રોટીનની અંદર સિસ્ટીન અવશેષો વચ્ચેના ડાયસલ્ફાઇડ બોન્ડની રચના અને તૂટવાનું ઉત્પ્રેરક કરે છે. PDI ને હાઉસકીપિંગ એન્ઝાઇમ ERO1 ની ક્રિયા દ્વારા રિસાયકલ કરવામાં આવે છે, જે PDI [41] માં ડિસલ્ફાઇડ બોન્ડને ફરીથી દાખલ કરે છે. મોલેક્યુલર ઓક્સિજન એ ERO1 નું ટર્મિનલ ઇલેક્ટ્રોન સ્વીકારનાર છે, જે ઉત્પાદિત દરેક ડિસલ્ફાઇડ બોન્ડ માટે હાઇડ્રોજન પેરોક્સાઇડની સ્ટોઇકિયોમેટ્રિક માત્રા ઉત્પન્ન કરે છે [42]. Peroxidases (PRX4) અને glutathione peroxidases (GPX7 અને GPX8) એ ER માં હાઇડ્રોજન પેરોક્સાઇડ ઘટાડવા માટેના મુખ્ય ઉત્સેચકો છે. જ્યારે આ ઓક્સિડો-રિડક્ટિવ સિસ્ટમ યોગ્ય રીતે કામ કરતી નથી, ત્યારે ER માં મિસફોલ્ડ પ્રોટીનનો અસામાન્ય સંચય થાય છે અને અનફોલ્ડ પ્રોટીન રિસ્પોન્સ (UPR) નામના સંકેતોનો સમૂહ ER હોમિયોસ્ટેસિસ [43] ને પુનઃસ્થાપિત કરવા માટે સાયટોપ્લાઝમ અને ન્યુક્લિયસમાં પ્રસારિત થાય છે. યુકેરીયોટ્સમાં ER સ્ટ્રેસને સેન્સ કરવા માટે ત્રણ મેમ્બ્રેન-સંબંધિત પ્રોટીનની ઓળખ કરવામાં આવી છે: ટ્રાન્સક્રિપ્શન ફેક્ટર 6 (ATF6), સ્વાદુપિંડનું ER eIF2 સક્રિય કરવું? કિનેઝ (PERK, પણ ડબલ-સ્ટ્રેન્ડેડ આરએનએ-સક્રિયકૃત પ્રોટીન કિનેઝ-જેવા ER કિનેઝ), અને ઇનોસિટોલ-જરૂરી કિનાઝ1 (IRE1). દરેક સેન્સરનું લ્યુમિનલ ડોમેન ગ્લુકોઝ-રેગ્યુલેટેડ પ્રોટીન (GRP78/BIP) તરીકે ઓળખાતા 78 kDa ચેપરોન સાથે બંધાયેલું છે. ER સ્ટ્રેસ પર BIP વિખરાયેલા પ્રોટીનને જોડવા માટે અલગ થઈ જાય છે, જે ત્રણ સેન્સર્સના સક્રિયકરણ તરફ દોરી જાય છે [44].
NRF2 અને તેના હોમોલોગ NRF1, એન્ટીઑકિસડન્ટ પ્રતિભાવ સાથે પણ સંબંધિત છે, યુપીઆરને ન્યુક્લિયસમાં ટ્રાન્સડક્શનમાં ભાગ લે છે. NRF1 ના કિસ્સામાં, આ પ્રોટીન ER મેમ્બ્રેન પર સ્થિત છે અને ડિગ્લાયકોસિલેશન અથવા ક્લીવેજ પર પરમાણુ ટ્રાન્સલોકેશનમાંથી પસાર થાય છે. પછી, UPR સક્રિયકરણ NRF1 ની પ્રક્રિયા અને પરમાણુ કમ્પાર્ટમેન્ટમાં પરિણામી ટુકડાના પરમાણુ સંચય તરફ દોરી જાય છે. જો કે, આ NRF1 ટુકડાના ARE-સમાવતી જનીનોને ટ્રાન્સએક્ટિવેટ કરવાની ક્ષમતા હજુ પણ ચર્ચા હેઠળ છે [45].
ગ્લોવર-કટર અને સહકાર્યકરોએ સી. એલિગન્સ, SKN-2 ના NRF1 ઓર્થોલોગને અલગ-અલગ ER સ્ટ્રેસર્સ સાથે સક્રિય કરવાનું દર્શાવ્યું. SKN-1 અભિવ્યક્તિમાં વધારો IRE1 અથવા PERK વોર્મ ઓર્થોલોગ્સ [46] સહિત વિવિધ UPR મધ્યસ્થીઓ પર આધારિત હતો. PERK-ઉણપવાળા કોષોમાં, ક્ષતિગ્રસ્ત પ્રોટીન સંશ્લેષણ અંતર્જાત પેરોક્સાઇડ્સ અને અનુગામી એપોપ્ટોસિસ [47] ના સંચય તરફ દોરી જાય છે. આ પેરોક્સાઇડ્સથી ER ને સુરક્ષિત કરવા માટે PERK દ્વારા ઉપયોગમાં લેવાતું ઇફેક્ટર NRF2 હોઈ શકે છે, કારણ કે એવું નોંધવામાં આવ્યું છે કે PERK ફોસ્ફોરીલેટ્સ NRF2 ને Ser40 પર, આમ KEAP1 [31] દ્વારા તેના અધોગતિને અટકાવે છે. ASK1 નું ઇન્ડક્શન પણ IRE2 [1] ની TRAF48- મધ્યસ્થી કિનાઝ ક્રિયા દ્વારા આ માર્ગમાં ભૂમિકા ભજવે તેવી શક્યતા છે. NRF2 ના નિયમનમાં MAPK ની ભૂમિકા હજી પણ વિવાદાસ્પદ હોવા છતાં, તાજેતરમાં એવું સૂચવવામાં આવ્યું હતું કે IRE1-TRAF2-ASK1-JNK પાથવે NRF2 [49] (ફિગ. 1) ને સક્રિય કરી શકે છે. રસપ્રદ રીતે, સી. એલિગન્સ અને માનવ કોષોમાં, નવા પુરાવા સૂચવે છે કે તેના સક્રિયકરણ લૂપ પર IRE1 કિનેઝનું સિસ્ટીન સલ્ફેનિલેશન IRE1- મધ્યસ્થી યુપીઆરને અટકાવે છે અને NRF38 દ્વારા સંચાલિત p2 એન્ટીઑકિસડન્ટ પ્રતિભાવ શરૂ કરે છે. ડેટા સૂચવે છે કે IRE1 એ સાયટોપ્લાઝમિક સેન્ટિનલ તરીકે પ્રાચીન કાર્ય ધરાવે છે જે p38 અને NRF2 [50] ને સક્રિય કરે છે.
આકૃતિ 1 UPR દ્વારા NRF2 નું નિયમન. એન્ડોપ્લાઝમિક રેટિક્યુલમની અંદર અનફોલ્ડ અથવા મિસફોલ્ડ પ્રોટીનનું સંચય અનફોલ્ડ પ્રોટીન પ્રતિભાવ (UPR) શરૂ કરી શકે છે. પ્રથમ, ચેપરોન BIP એ ER સેન્સર IRE1 અને PERK ના ઇન્ટ્રાલ્યુમિનલ ડોમેનમાંથી અનફોલ્ડ/મીસફોલ્ડ પ્રોટીનને બાંધવા માટે મુક્ત કરવામાં આવે છે. આ તેમના સાયટોસોલિક ડોમેન્સના ડાયમરાઇઝેશન અને ટ્રાન્સ-ઓટો-ફોસ્ફોરાયલેશનને સક્ષમ કરે છે. PERK સક્રિયકરણ Ser2 પર સીધા NRF40 ફોસ્ફોરાયલેશનમાં પરિણમે છે, જે NRF2 ને ન્યુક્લિયસમાં સ્થાનાંતરણ તરફ દોરી જાય છે અને લક્ષ્ય જનીનો સક્રિય થાય છે. IRE1 સક્રિયકરણ TRAF2 ની ભરતીને પ્રેરિત કરે છે ત્યારબાદ ASK1 અને JNK ફોસ્ફોરાયલેશન અને સક્રિયકરણ. જેમ કે JNK ને ફોસ્ફોરીલેટ અને NRF2 ને સક્રિય કરવાની જાણ કરવામાં આવી છે, તે વિચારવું વાજબી છે કે IRE1 સક્રિયકરણ NRF2 પ્રવૃત્તિમાં વધારો કરશે.
પ્રોટીન ગ્લાયકોસિલેશન ટ્યુનિકામિસિનના અવરોધક સાથે UPR ના ઇન્ડક્શન પર ઘણા અભ્યાસ હાથ ધરવામાં આવ્યા છે. NRF2 ટ્યુનિકામિસિન-પ્રેરિત એપોપ્ટોટિક કોષોના મૃત્યુને રોકવા માટે આવશ્યક જણાય છે [31] અને આ પરિસ્થિતિઓ હેઠળ તેનું સક્રિયકરણ KEAP1 [51] ના ઓટોફેજિક ડિગ્રેડેશન દ્વારા સંચાલિત થાય છે. તદનુસાર, ?TC-2 કોષોમાં NRF6 અભિવ્યક્તિનું shRNA-મધ્યસ્થી મૌન, એક મ્યુરિન ઇન્સ્યુલિનોમા?-સેલ લાઇન, નોંધપાત્ર રીતે ટ્યુનિકામિસિન-પ્રેરિત સાયટોટોક્સિસિટીમાં વધારો થયો અને પ્રો-એપોપ્ટોટિક ER સ્ટ્રેસ માર્કર CHOP10 ની અભિવ્યક્તિમાં વધારો થયો. બીજી બાજુ, 2-ડિથિઓલ-1,2-થિઓન (D3T) દ્વારા NRF3 સક્રિયકરણે ટ્યુનિકામિસિન સાયટોટોક્સિસિટીમાં ઘટાડો કર્યો અને CHOP10 અને PERK [52] ની અભિવ્યક્તિને ઓછી કરી. રસપ્રદ વાત એ છે કે, ઘ્રાણેન્દ્રિયને લગતું ચેતાકોષો ટ્યુનિકામિસિનના પ્રણાલીગત ઉપયોગને સબમિટ કરે છે અને અન્ય યુપીઆર-સભ્યો જેમ કે CHOP, BIP, XBP2 [1] સાથે સમાંતર NRF53 માં વધારો કરે છે. આ પરિણામોને વિવો અભ્યાસમાં વિસ્તારવામાં આવ્યા છે, કારણ કે ઉંદરોમાં ટ્યુનિકામિસિનનું લેટરલ વેન્ટ્રિક્યુલર ઇન્ફ્યુઝન હિપ્પોકેમ્પસમાં PERK અને NRF2 ની અભિવ્યક્તિને પ્રેરિત કરે છે, જેની સાથે નોંધપાત્ર જ્ઞાનાત્મક ખામીઓ, TAU ફોસ્ફોરીલેશન અને A?42 ડિપોઝિટમાં વધારો થાય છે [54].
NRF2 ER ફિઝિયોલોજીના જાળવણી માટે મુખ્ય જનીનો ઉપર-નિયમન કરે છે
ER લ્યુમેનને ડાયસલ્ફાઇડ રસાયણશાસ્ત્ર જાળવવા માટે સાયટોસોલમાંથી GSH ના પુષ્કળ પુરવઠાની જરૂર છે. NRF2 મગજમાં GSH ચયાપચયના નિર્ણાયક ઉત્સેચકોને મોડ્યુલેટ કરે છે, જેમ કે સિસ્ટીન/ગ્લુટામેટ ટ્રાન્સપોર્ટ, ?-ગ્લુટામેટ સિસ્ટીન સિન્થેટેઝ (?-GS), ગ્લુટામેટ-સિસ્ટીન લિગેસ કેટાલિટીક અને મોડ્યુલેટર સબ્યુનિટ્સ (GCLC અને GCLM), ગ્લુટાથિઓન (GCLC અને GCLM) glutathione peroxidase (GPX) ([55] માં સમીક્ષા). ER માં GSH ની જાળવણીમાં NRF2 ની સુસંગતતા એ શોધ દ્વારા સમર્થિત છે કે NRF2 નું ફાર્માકોલોજિકલ અથવા આનુવંશિક સક્રિયકરણ GCLC/GCLM દ્વારા GSH સંશ્લેષણમાં પરિણમે છે, જ્યારે NRF2-નોકડાઉન દ્વારા આ ઉત્સેચકોની અભિવ્યક્તિને અટકાવવાથી નુકસાનનું સંચય થાય છે. ER ની અંદર પ્રોટીન UPR સક્રિયકરણ [56] તરફ દોરી જાય છે.
C. માં SKN-1 દ્વારા નિયમન કરાયેલ UPR લક્ષ્ય જનીનોના ઘણા ઘટકો, જેમાં Ire1, Xbp1 અને Atf6 નો સમાવેશ થાય છે. જો કે NRF2 સસ્તન પ્રાણીઓમાં ઘણા પેરોક્સિડેઝ (PRX) અને ગ્લુટાથિઓન પેરોક્સિડેઝ (GPX) જનીનોની અભિવ્યક્તિનું નિયમન કરે છે ([57] માં સમીક્ષા કરવામાં આવી છે), માત્ર GPX8 એ એક પ્રામાણિક ER-સ્થાનિક એન્ઝાઇમ છે, જે KDEL પુનઃપ્રાપ્તિ સંકેત [58] ને આશ્રય આપે છે. GPX8 ની ખોટ યુપીઆર સક્રિયકરણ, ERO1 નું લીકેજ?-ઉત્પાદિત હાઇડ્રોજન પેરોક્સાઇડ સાયટોસોલ અને કોષ મૃત્યુનું કારણ બને છે. હાઇડ્રોજન પેરોક્સાઇડ ERO1 માંથી મેળવવામાં આવે છે? GPX8 અને PRX4 [59] ની સંકલિત ક્રિયાને કારણે પ્રવૃત્તિ ER થી સાયટોસોલ સુધી ફેલાઈ શકતી નથી. આ સંદર્ભમાં, જંગલી પ્રકાર અને NRF2-નલ ઉંદર પેશીઓમાંથી આરએનએનો ઉપયોગ કરીને એન્ટીઑકિસડન્ટ સંરક્ષણ પાથવે-જીન અભિવ્યક્તિ એરેનું વિશ્લેષણ દર્શાવે છે કે NRF8 [2] ની ગેરહાજરીમાં GPX60 ની અભિવ્યક્તિ ડાઉન-રેગ્યુલેટેડ હતી. આને અનુરૂપ, માયલોપ્રોલિફેરેટિવ નિયોપ્લાઝમ, પોલિસિથેમિયા અથવા માયલોફિબ્રોસિસથી પીડિત દર્દીના નમૂનાઓમાંથી ટ્રાન્સક્રિપ્ટોમ વિશ્લેષણ, રોગો પણ ઓક્સિડેટીવ તણાવ અને લો-ગ્રેડ ક્રોનિક ઇન્ફ્લેમેશન સાથે સંકળાયેલા છે, નિયંત્રણ વિષયોની સરખામણીમાં NRF2 અને GPX8 બંનેના નીચા અભિવ્યક્તિ સ્તર દર્શાવે છે [61]. હજી સુધી એવા અભ્યાસો નથી કે જેમાં ખાસ કરીને માનવ મગજની સુરક્ષામાં GPX8 સામેલ હોય પરંતુ ઉંદરમાં ટ્રાન્સક્રિપ્ટમ વિશ્લેષણ પાર્કિન્સોનિયન ટોક્સિન MPTP [8] ના પ્રતિભાવમાં વળતર આપનાર GPX62 વધારો દર્શાવે છે.
ન્યુરોડિજનરેટિવ રોગોમાં યુપીઆર ડિસરેગ્યુલેશન પર NRF2 ની અસર
પીડીઆઈ ઉત્સેચકોની ખામી અને યુપીઆરનું ક્રોનિક સક્રિયકરણ પછીથી ન્યુરોડિજનરેશન શરૂ અથવા વેગ આપી શકે છે. રોગ-અસરગ્રસ્ત ચેતાકોષો, ન્યુરોડિજનરેટિવ રોગના પ્રાણી મોડેલો તેમજ પોસ્ટ-મોર્ટમ માનવ પેશીઓ આમાંના મોટા ભાગના વિકારોમાં કેટલાક UPR-માર્કર્સના અપ-નિયમનના પુરાવા આપે છે. ન્યુરોડિજનરેટિવ રોગોમાં PDI/UPR પાથવેના ફેરફારની [63] માં સરસ રીતે સમીક્ષા કરવામાં આવી છે પરંતુ મગજના પોસ્ટમોર્ટમ નમૂનાઓમાંથી નીચેના હાઇલાઇટ્સને ધ્યાનમાં લેવા જોઈએ. ગૂંચવાયેલા ચેતાકોષોમાં અને એડી અને પીડી દર્દીઓના લેવી બોડીઝમાં અનુક્રમે PDI સ્તર વધે છે [64], [65]. PDI અને ERP57 CSF માં ALS દર્દીઓ અને CJD વિષયો [66], [67], [68] ના મગજમાં ઉપર-નિયમિત છે. AD, PD અથવા ALS [1], [6], [69], [70] ધરાવતા દર્દીઓના નમૂનાઓમાં BIP, PERK, IRE71 અને ATF67 એ એલિવેટેડ છે. એચડી [1], [72] માંથી પોસ્ટમોર્ટમ મગજના નમૂનાઓમાં BIP, CHOP અને XBP73 ઊંચા છે. વધુમાં, ERP57, GRP94 અને BIP નું અપ-રેગ્યુલેશન CJD દર્દીઓ [74] ના કોર્ટેક્સ પેશીઓમાં જોવા મળ્યું હતું. એકંદરે, આ પુરાવા દર્શાવે છે કે મગજના પેરેન્ચાઇમામાં ખોટા ફોલ્ડ પ્રોટીનનું સંચય યુપીઆરના નુકસાનકારક અને ક્રોનિક સક્રિયકરણ તરફ દોરી જાય છે. રસપ્રદ વાત એ છે કે, AD ની શરૂઆતમાં PERK દ્વારા NRF2 ના સક્રિયકરણને જોડતો એક તાજેતરનો અભ્યાસ છે. આ અભ્યાસમાં, લેખકોએ વિશ્લેષણ કર્યું કે શું ઓક્સિડેટીવ તણાવ NRF2 અને UPR માં મધ્યસ્થી ફેરફારો માનવ પેરિફેરલ રક્ત કોશિકાઓ અને એડી ટ્રાન્સજેનિક માઉસ મોડેલનો ઉપયોગ કરીને એડી પેથોજેનેસિસમાં પ્રારંભિક ઘટનાઓનું નિર્માણ કરી શકે છે કે કેમ તે વિવિધ રોગના તબક્કામાં. માનવ પેરિફેરલ બ્લડ મોનોન્યુક્લિયર કોષોમાં ઓક્સિડેટીવ તણાવમાં વધારો અને pSer40-NRF2 માં વધારો જોવા મળ્યો હતો જે હળવી જ્ઞાનાત્મક ક્ષતિ ધરાવતા વ્યક્તિઓથી અલગ પડે છે. વધુમાં, તેઓએ હળવી જ્ઞાનાત્મક ક્ષતિ અને હળવી એડી [75] ધરાવતા વ્યક્તિઓમાંથી આ કોષોમાં અશક્ત ER કેલ્શિયમ હોમિયોસ્ટેસિસ અને અપ-રેગ્યુલેટેડ ER-સ્ટ્રેસ માર્કર્સની જાણ કરી.
NRF2 અને યુબીક્વિટીન પ્રોટીઝોમ સિસ્ટમ (યુપીએસ)નું મ્યુચ્યુઅલ રેગ્યુલેશન
UPS NRF2 પ્રોટીન સ્તરોને મોડ્યુલેટ કરે છે
યુપીએસ ક્ષતિગ્રસ્ત અથવા ખોટા ફોલ્ડ પ્રોટીનના અધોગતિમાં ભાગ લે છે અને સાયટોસોલ અને ન્યુક્લિયસમાં મુખ્ય નિયમનકારી અણુઓના સ્તરને નિયંત્રિત કરે છે. આ સિસ્ટમનો કેન્દ્રિય કોર એક વિશાળ મલ્ટિસુબ્યુનિટ એન્ઝાઇમ છે જેમાં 20S નામનું પ્રોટીઓલિટીક સક્રિય સંકુલ છે. 20S કોર પ્રોટીઝોમ અનફોલ્ડ પ્રોટીનને ડિગ્રેડ કરે છે, પરંતુ વિવિધ રેગ્યુલેટરી પ્રોટીન કોમ્પ્લેક્સ સાથે જોડાવાથી તેની સબસ્ટ્રેટની વિશિષ્ટતા અને પ્રવૃત્તિ બદલાય છે. દાખલા તરીકે, 19S કોરમાં એક કે બે 20S રેગ્યુલેટરી સબ્યુનિટ્સનો ઉમેરો 26S પ્રોટીઝોમ બનાવે છે અને મૂળ ફોલ્ડ પ્રોટીન [76], [77] તરફ તેની વિશિષ્ટતાને બદલે છે. પ્રોટીસોમલ ડિગ્રેડેશન માટે યુબીક્વિટીનનું સહસંયોજક બંધન જરૂરી છે. યુબીક્વિટીનનું જોડાણ ત્રણ-પગલાની કાસ્કેડ પદ્ધતિ દ્વારા આગળ વધે છે. પ્રથમ, યુબીક્વિટીન-સક્રિય કરનાર એન્ઝાઇમ E1 એટીપી-જરૂરી પ્રતિક્રિયામાં યુબીક્વિટીનને સક્રિય કરે છે. પછી, એક E2 એન્ઝાઇમ (યુબીક્વિટીન-કેરિયર પ્રોટીન અથવા યુબીક્વિટીન-કન્જુગેટીંગ એન્ઝાઇમ) સક્રિય યુબીક્વિટીનને E1 થી સબસ્ટ્રેટમાં સ્થાનાંતરિત કરે છે જે ખાસ કરીને યુબીક્વિટીન-પ્રોટીન લિગેઝ પરિવારના સભ્ય સાથે બંધાયેલ છે, જેનું નામ E3 છે. જોકે સર્વવ્યાપક-પ્રોટીનનું ચોક્કસ ભાવિ યુબીક્વિટીન સાંકળની પ્રકૃતિ પર આધારિત હશે, આ પ્રક્રિયા સામાન્ય રીતે 26S પ્રોટીઝોમ [78] દ્વારા અધોગતિમાં પરિણમે છે.
E3-ligase KEAP1 એ NRF2 નું સૌથી જાણીતું અવરોધક છે. KEAP1 નિયમનની પદ્ધતિ સુંદર રીતે સમજાવે છે કે NRF2 સ્તર ઓક્સિડન્ટ વધઘટ સાથે કેવી રીતે સમાયોજિત થાય છે. મૂળભૂત સ્થિતિમાં, નવા સંશ્લેષિત NRF2 ને હોમોડીમર KEAP1 દ્વારા પકડવામાં આવે છે, જે એક NRF2 પરમાણુને બે એમિનો એસિડ સિક્વન્સ પર નીચા (એસ્પાર્ટેટ, લ્યુસીન, ગ્લાયસીન; DLG) અને ઉચ્ચ (ગ્લુટામેટ, થ્રેઓનિન, ગ્લુટામેટ, થ્રેઓનિન, ગ્લુટામેટ, ગ્લુટામેટ) સાથે જોડે છે. KEAP1 સાથેની ક્રિયાપ્રતિક્રિયા NRF2 ને CULLIN3/RBX1 પ્રોટીન સંકુલમાં રજૂ કરવામાં મદદ કરે છે, પરિણામે તેનું સર્વવ્યાપકીકરણ અને અનુગામી પ્રોટીસોમલ ડિગ્રેડેશન થાય છે. જો કે, KEAP1 નું રેડોક્સ ફેરફાર CULLIN2/RBX3 દ્વારા રજૂ કરાયેલ UPS પર NRF1 ની રજૂઆતને અવરોધે છે. પરિણામે, નવા સંશ્લેષિત NRF2 KEAP1-આશ્રિત અધોગતિથી બચી જાય છે, ન્યુક્લિયસમાં એકઠા થાય છે અને ARE-સમાવતી જનીનોને સક્રિય કરે છે [79], [80], [81], [82].
E3-ligase એડેપ્ટર?-TrCP એ હોમોડીમર પણ છે જે GSK-2 દ્વારા NRF3 ના ફોસ્ફોરાયલેશન સંબંધિત સિગ્નલિંગ ઇવેન્ટ્સમાં ભાગ લે છે. આ કિનાઝ ફોસ્ફોરીલેટ્સ NRF2 (એસ્પાર્ટેટ, સેરીન, ગ્લાયસીન, આઇસોલ્યુસીન સેરીન; DSGIS) ના ચોક્કસ સેરીન અવશેષોને ડિગ્રેડેશન ડોમેન બનાવવા માટે બનાવે છે જે પછી ?-TrCP દ્વારા ઓળખાય છે અને CULLIN1/RBX1 કોમ્પ્લેક્સ દ્વારા પ્રોટીઝોમ ડિગ્રેડેશન માટે ટૅગ કરવામાં આવે છે. GSK-3 દ્વારા ફોસ્ફોરીલેટેડ ચોક્કસ એમિનો એસિડની ઓળખ? આ ડીગ્રોનમાં Neh6 ડોમેન, 2D-જેલ ઇલેક્ટ્રોફોરેસીસ [15], [26] અને માસ સ્પેક્ટ્રોસ્કોપી [83] ના સાઇટ-નિર્દેશિત મ્યુટાજેનેસિસના સંયોજન દ્વારા હાથ ધરવામાં આવ્યું હતું. પરિણામે, GSK-3 નું નિષેધ? GSK-3 isoforms સામે અત્યંત પસંદગીયુક્ત દવાઓ અથવા siRNAs દ્વારા NRF2 પ્રોટીન સ્તરમાં વધારો થયો છે. ?-TrCP આઇસોફોર્મ્સ 1 અને 2 સામે siRNAs સાથે સમાન પરિણામો જોવા મળ્યા હતા. GSK-2 ને અનુસરતા NRF3 નું સ્થિરીકરણ? KEAP1-ની ઉણપ ધરાવતા માઉસ એમ્બ્રીયો ફાઈબ્રોબ્લાસ્ટ્સમાં અને KEAP2-સ્વતંત્ર નિયમનનું વધુ નિદર્શન કરતાં, KEAP1 સાથે ઉચ્ચ-સંબંધ બંધનકર્તા માટે નિર્ણાયક ETGE અવશેષોનો અભાવ ધરાવતા એક્ટોપિકલી વ્યક્ત NRF1 ડિલીશન મ્યુટન્ટમાં અવરોધ જોવા મળ્યો હતો.
ન્યુરોડિજનરેટિવ રોગોના સંદર્ભમાં, અમે બે અલગ અલગ રીતે UPS દ્વારા NRF2 ના મોડ્યુલેશનની કલ્પના કરી શકીએ છીએ. એક તરફ, KEAP1 સિસ્ટમ મિસફોલ્ડ પ્રોટીન સંચયમાંથી મેળવેલા રેડોક્સ અસંતુલનને સમજશે, જ્યારે GSK-3/?-TrCP અક્ષ પ્રોટીઓસ્ટેસિસ (ફિગ. 2) ના નુકશાન દ્વારા બદલાયેલ સિગ્નલિંગ ટ્રાન્સડક્શનમાં સક્રિય સહભાગી તરીકે કાર્ય કરશે.
આકૃતિ 2 UPS NRF2 સ્તરને ચુસ્તપણે નિયંત્રિત કરે છે. હોમિયોસ્ટેટિક પરિસ્થિતિઓમાં, નીચા NRF2 સ્તરને E3 લિગેસેસ એડેપ્ટરો KEAP1 અને ?-TrCP ની ક્રિયા દ્વારા જાળવવામાં આવે છે. ડાબે, NRF2 નીચા (DLG) અને ઉચ્ચ (ETGE) એફિનિટી મોટિફ્સ દ્વારા KEAP1 હોમોડિમરના કેલ્ચ ડોમેન્સ સાથે જોડાય છે. તેના BTB ડોમેન દ્વારા, KEAP1 એક સાથે CULLIN3/RBX1 સંકુલ સાથે જોડાય છે, જે NRF2 સર્વવ્યાપકતા અને 26 S પ્રોટીઝોમ દ્વારા અધોગતિને સક્ષમ કરે છે. વધુમાં, GSK-3? ડિગ્રેડેશન ડોમેન (DpSGIpSL) બનાવવા માટે NRF335 ના ફોસ્ફોરીલેટ્સ Ser338 અને Ser2 અવશેષો કે જે પછી ubiquitin ligase એડેપ્ટર ?-TrCP દ્વારા ઓળખાય છે અને CULLIN3/RBX1 કોમ્પ્લેક્સ દ્વારા પ્રોટીઝોમ ડિગ્રેડેશન માટે ટેગ કરવામાં આવે છે. જમણે, પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓ અથવા ઇલેક્ટ્રોફાઇલ્સના સંપર્કમાં આવવા પર KEAP1 માં ગંભીર Cys અવશેષો સંશોધિત કરવામાં આવે છે, જે KEAP1 ને NRF2 અથવા CULLIN3/RBX1 સાથે કાર્યક્ષમ રીતે ક્રિયાપ્રતિક્રિયા કરવામાં અસમર્થ બનાવે છે અને પછી આ ટ્રાન્સક્રિપ્શન પરિબળ તેની અર્ધ-જીવન અને ટ્રાન્સક્રિપ્શનલ પ્રવૃત્તિમાં વધારો કરે છે. GSK-3?, જેમ કે Ser9 ખાતે AKT ફોસ્ફોરાયલેશનના નિષેધમાં પરિણમે છે તેવા સંકેતોના માર્ગો, પ્રોટીઝોમ, સંચય અને લક્ષ્ય જનીનો ઇન્ડક્શન દ્વારા NRF2 ક્ષતિગ્રસ્ત અધોગતિમાં પરિણમે છે.
NRF2 પ્રોટીઝોમ સબ્યુનિટ્સના ટ્રાન્સક્રિપ્શનલ કંટ્રોલ દ્વારા UPS પ્રવૃત્તિમાં વધારો કરે છે
NRF2 ઘણા પ્રોટીઝોમ સબ્યુનિટ્સની અભિવ્યક્તિનું નિયમન કરે છે, આમ કોષને ઝેરી પ્રોટીનના સંચયથી રક્ષણ આપે છે. NRF2 ઇન્ડ્યુસર D2T [3] સાથે સુયોજિત લિવર આરએનએના વિશાળ માઇક્રોએરે વિશ્લેષણ અનુસાર, વીસ પ્રોટીઝોમ- અને સર્વવ્યાપકતા-સંબંધિત જનીનો NRF84 દ્વારા નિયમન કરવામાં આવે છે. પશ્ચાદવર્તી અભ્યાસમાં, સમાન લેખકોએ પુરાવો આપ્યો હતો કે 26S પ્રોટીઝોમના મોટાભાગના સબ્યુનિટ્સની અભિવ્યક્તિ D3T સાથે સારવાર કરાયેલા ઉંદરોના યકૃતમાં ત્રણ ગણી સુધી વધારી હતી. સબ્યુનિટ પ્રોટીનનું સ્તર અને પ્રોટીઝોમ પ્રવૃત્તિ સંકલનપૂર્વક વધારવામાં આવી હતી. જો કે, જ્યાં ટ્રાન્સક્રિપ્શન ફેક્ટર NRF2 વિક્ષેપિત થયું હોય ત્યાં ઉંદરમાં કોઈ ઇન્ડક્શન જોવા મળ્યું ન હતું. PSMB5 (20S) પ્રોટીઝોમ સબ્યુનિટની પ્રમોટર પ્રવૃત્તિ NRF2 ઓવરએક્સપ્રેશન અથવા માઉસ એમ્બ્રીયોનિક ફાઈબ્રોબ્લાસ્ટ્સમાં એક્ટિવેટર્સ સાથેની સારવાર સાથે વધી હતી, અને PSMB5 [85] ના પ્રોક્સિમલ પ્રમોટરમાં AREs ઓળખવામાં આવ્યા હતા. NRF2 ના ફાર્માકોલોજિકલ સક્રિયકરણના પરિણામે પ્રતિનિધિ પ્રોટીઝોમ સબ્યુનિટ્સ (PSMA3, PSMA6, PSMB1 અને PSMB5) ના ઉચ્ચ અભિવ્યક્તિ સ્તરો માત્ર કાર્યાત્મક NRF2 [86] ધરાવતા બિન-સંવેદનશીલ માનવ ફાઇબ્રોબ્લાસ્ટ્સમાં પરિણમ્યા. ઓક્સિડેટીવ તણાવમાં અનુકૂલન દરમિયાન NRF2 સક્રિયકરણ PSMB1 (20S) અને PA28 ની ઉચ્ચ અભિવ્યક્તિમાં પરિણમે છે? સબયુનિટ્સ (અથવા S11, પ્રોટીઝોમ રેગ્યુલેટર) [87]. વધુમાં, માનવ ગર્ભના સ્ટેમ કોશિકાઓના પરિણામો પરથી જાણવા મળ્યું કે NRF2 પ્રોટીઝોમ પરિપક્વતા પ્રોટીન (POMP) ની અભિવ્યક્તિને નિયંત્રિત કરે છે, એક પ્રોટીઝોમ ચેપરોન, જે બદલામાં સ્વ-નવીકરણ માનવ ગર્ભ સ્ટેમ કોશિકાઓના પ્રસારને, ત્રણ જર્મ લેયર ડિફરન્સિએશન અને સેલ્યુલર રિપ્રોગ્રામિંગ [ 88]. બધા એકસાથે, આ અભ્યાસો સૂચવે છે કે NRF2 UPS ના મુખ્ય ઘટકોની અભિવ્યક્તિનું નિયમન કરે છે અને તેથી પ્રોટીનની મંજૂરીમાં સક્રિયપણે ફાળો આપે છે જે અન્યથા ઝેરી હશે.
ન્યુરોડિજનરેટિવ રોગોમાં NRF2-UPS એક્સિસ
ન્યુરોડિજનરેટિવ રોગોમાં યુપીએસની ભૂમિકા સઘન ચર્ચાનું ક્ષેત્ર છે. પ્રારંભિક અભ્યાસોએ અનેક ન્યુરોડિજનરેટિવ રોગોથી અસરગ્રસ્ત દર્દીઓની માનવ નેક્રોપ્સીમાં પ્રોટીઝોમ પ્રવૃત્તિમાં ઘટાડો નોંધ્યો હતો. જો કે, વિટ્રો અને વિવો અભિગમમાં રોજગારી આપતા અન્ય અભ્યાસોમાં અપરિવર્તિત અથવા તો પ્રોટીઝોમ પ્રવૃત્તિમાં વધારો જોવા મળ્યો છે ([89] માં સમીક્ષા કરવામાં આવી છે). આ વિસંગતતા માટે એક સંભવિત સમજૂતી એ છે કે NRF2-લક્ષ્યો માટે સૂચવવામાં આવ્યું છે તેમ રોગની પ્રગતિ દરમિયાન અને મગજના વિવિધ પ્રદેશોમાં UPS ઘટકોના સ્તરો બદલાઈ શકે છે.
આ વિવાદ હોવા છતાં, એ નોંધવું જોઈએ કે ARE-સમાવતી પ્રોટીઝોમ જનીનોનું અપ-રેગ્યુલેશન મગજમાં ઝેરી પ્રોટીનના ક્લિયરન્સને વધારીને UPSને મજબૂત બનાવશે. ખરેખર, ન્યુરોનલ કોશિકાઓમાં એન્ટીઑકિસડન્ટ પ્રતિભાવના મોડ્યુલેટર, NRF1 નું નિરાકરણ ક્ષતિગ્રસ્ત પ્રોટીઝોમ પ્રવૃત્તિ અને ન્યુરોડિજનરેશન તરફ દોરી જાય છે. ક્રોમેટિન ઇમ્યુનોપ્રિસિપિટેશન પ્રયોગો અને ટ્રાન્સક્રિપ્શનલ વિશ્લેષણ દર્શાવે છે કે PSMB6 NRF1 દ્વારા નિયંત્રિત થાય છે. વધુમાં, જનીન અભિવ્યક્તિ રૂપરેખાને કારણે NRF1 ને ચેતાકોષોમાં પ્રોટીઝોમ જનીનોના મુખ્ય ટ્રાન્સક્રિપ્શનલ રેગ્યુલેટર તરીકે ઓળખવામાં આવી, જે સૂચવે છે કે NRF1 માં વિક્ષેપ ન્યુરોડિજનરેટિવ રોગોના પેથોજેનેસિસમાં ફાળો આપી શકે છે [90]. રસપ્રદ રીતે, NRF1 અને તેના લાંબા આઇસોફોર્મ જેને TCF11 કહેવાય છે તે પ્રોટીસોમ અવરોધ પર ARE-સમાવતી પ્રોટીઝોમ જનીનોને પ્રતિસાદ લૂપમાં ઘટાડેલી પ્રોટીઓલિટીક પ્રવૃત્તિને વળતર આપવા માટે દર્શાવવામાં આવ્યું હતું
NRF2 વિશે, ન્યુરોટોક્સિન પેરાક્વેટ [2] સાથે સારવાર કરાયેલ ડીજે-6-ઉણપવાળા ઉંદરના મધ્ય મગજમાં NRF19, RPT5 (20 S) અને PSMB1 (93 S) સ્તરના ઘટાડા વચ્ચે સહસંબંધ છે. વધુમાં, કુદરતી રીતે બનતું સંયોજન સલ્ફોરાફેન (SFN) UPS ના નિર્ણાયક મોડ્યુલેટર તરીકે NRF2 ની વધુ મજબૂત છબી આપે છે. મ્યુરિન ન્યુરોબ્લાસ્ટોમા ન્યુરો2એ કોશિકાઓ સાથેના વિટ્રો પ્રયોગોએ પ્રોટીઝોમના ઉત્પ્રેરક સબ્યુનિટ્સ તેમજ SFN ના પ્રતિભાવમાં તેની પેપ્ટીડેઝ પ્રવૃત્તિઓની ઉન્નત અભિવ્યક્તિનો પુરાવો આપ્યો. આ દવા કોષોને હાઇડ્રોજન પેરોક્સાઇડ-મધ્યસ્થી સાયટોટોક્સિસિટી અને પ્રોટીન ઓક્સિડેશનથી પ્રોટીઝોમ ફંક્શન [94] પર આધારિત રીતે સુરક્ષિત કરે છે. વધુમાં, લિયુ અને સહકાર્યકરોએ મગજમાં SFN ના પ્રતિભાવમાં UPS પ્રવૃત્તિ પર દેખરેખ રાખવા માટે એક રિપોર્ટર માઉસનો ઉપયોગ કર્યો. આ ઉંદર સર્વવ્યાપક રીતે ગ્રીન ફ્લોરોસેન્સ પ્રોટીન (GFP) ને એક રચનાત્મક ડિગ્રેડેશન સિગ્નલ સાથે જોડે છે જે UPS (GFPu) દ્વારા તેના ઝડપી અધોગતિને પ્રોત્સાહન આપે છે. સેરેબ્રલ કોર્ટેક્સમાં, SFN એ 5 S પ્રોટીઝોમની કાયમોટ્રીપ્સિન જેવી (PSMB2), કેસ્પેસ જેવી (PSMB1), અને ટ્રિપ્સિન જેવી (PSMB20) પ્રવૃત્તિઓમાં સમાંતર વધારા સાથે GFPu નું સ્તર ઘટાડ્યું. વધુમાં, SFN સાથે હંટીંગ્ટન-પ્રાપ્ત કોષોની સારવારથી જાણવા મળ્યું છે કે NRF2 સક્રિયકરણ mHtt અધોગતિને વધારે છે અને mHtt સાયટોટોક્સિસિટી [95] ઘટાડે છે. SFN ક્રિયાની મુખ્ય પદ્ધતિ NRF2 [96] ના ઇન્ડક્શન દ્વારા છે. આગળના અભ્યાસોમાં NRF2-નલ સિસ્ટમનો ઉપયોગ કરીને NRF2 ના વિશિષ્ટ યોગદાનને ધ્યાનમાં લેવું જોઈએ.
NRF2 અને Macroautophagy વચ્ચે કાર્યાત્મક જોડાણ
NRF2 પ્રોટીન સ્તરો એડેપ્ટર પ્રોટીન P62 દ્વારા મોડ્યુલેટ કરવામાં આવે છે
ઓટોફેજી એ લિસોસોમની અંદર સાયટોસોલિક ઘટકોના અધોગતિનો ઉલ્લેખ કરે છે. આ પ્રક્રિયાનો ઉપયોગ લાંબા ગાળાના અને ખોટા ફોલ્ડ પ્રોટીન તેમજ ક્ષતિગ્રસ્ત ઓર્ગેનેલ્સના ક્લિયરન્સ માટે થાય છે. NRF2 અને ઓટોફેજી વચ્ચેનો સીધો સંબંધ સૌપ્રથમ એડેપ્ટર પ્રોટીન p62ના સંબંધમાં જોવા મળ્યો હતો, જેને SQSTM1 [97], [98], [99], [100], [101] પણ કહેવાય છે. આ પ્રોટીન પ્રોટીસોમલ અને લિસોસોમલ ડિગ્રેડેશન મશીનરીમાં સર્વવ્યાપક પ્રોટીનને શટલ કરે છે અને ક્ષતિગ્રસ્ત પ્રોટીનને તેમના અધોગતિ પહેલા એકંદરમાં અલગ પાડે છે. P62 સર્વવ્યાપક પ્રોટીનને બંધનકર્તા માટે, એક ubiquitin-સંબંધિત (UBA) ડોમેન રજૂ કરે છે, અને ઓટોફેજી રીસેપ્ટર LC3 દ્વારા ઓટોફેગોસોમલ મેમ્બ્રેન સાથે એકીકરણ માટે LC3-ઇન્ટરેક્ટિંગ રિજન (LIR) રજૂ કરે છે.
NRF62 અને તેના લક્ષ્ય જનીનોનું p2-મધ્યસ્થી ઇન્ડક્શન સૌપ્રથમ 2007 [102] માં નોંધાયું હોવા છતાં, KEAP1 [103], [98], [99], [100] સાથે તેની ક્રિયાપ્રતિક્રિયાની શોધ ન થાય ત્યાં સુધી મોલેક્યુલર મિકેનિઝમ સંપૂર્ણપણે સમજી શકાયું ન હતું. ], [101]. કોમાત્સુ અને સહકાર્યકરોએ p1 માં KEAP62 ઇન્ટરેક્ટિંગ રિજન (KIR) ને ઓળખ્યા જે KEAP1 ને NRF2 જેવા જ મૂળભૂત સપાટીના ખિસ્સામાં બાંધે છે અને NRF2 માં ETGE મોટિફ જેવું બંધનકર્તા જોડાણ ધરાવે છે, જે p62 અને NRF2 વચ્ચે સ્પર્ધાનું સૂચન કરે છે. p351 (62-DPSTGE-349) માં KIR મોટિફમાં Ser354 નું ફોસ્ફોરાયલેશન NRF1 બંધનકર્તા સાથે સ્પર્ધા કરીને અને તેના લક્ષ્ય જનીન [2], [98] ના સંચય અને ટ્રાન્સક્રિપ્શનલ સક્રિયકરણને મંજૂરી આપતા, KEAP99 માટે તેની આકર્ષણને વધારવા માટે દર્શાવવામાં આવ્યું હતું. વાસ્તવમાં, p62 ઓવરએક્સપ્રેશનને કારણે NRF2 સર્વવ્યાપકતામાં ઘટાડો થયો અને તેના પરિણામે સ્થિરીકરણ તેમજ તેના લક્ષ્ય જનીનોનું ઇન્ડક્શન થયું [104]. કેટલાક કિનાસને p62 ફોસ્ફોરીલેશનમાં ભાગ લેવાનું સૂચન કરવામાં આવ્યું છે. રેપામિસિન કોમ્પ્લેક્સ 1 (mTORC1) ના સસ્તન પ્રાણીઓના લક્ષ્યને સામેલ કરી શકાય છે, કારણ કે mTOR અવરોધક રેપામિસિન સાથેની સારવારથી p62 ના ફોસ્ફોરીલેશન અને આર્સેનાઇટ સારવાર પર KEAP1 ના ડાઉન-રેગ્યુલેશનને દબાવી દેવામાં આવે છે. તાજેતરમાં, એવું દર્શાવવામાં આવ્યું હતું કે TGF-?-activated kinase 1 (TAK1) p62 ફોસ્ફોરીલેટ પણ કરી શકે છે, KEAP1 ડિગ્રેડેશન અને NRF2-અપ-રેગ્યુલેશનને વધારે છે. આ અભ્યાસના લેખકો સૂચવે છે કે સ્થિર-સ્થિતિમાં સેલ્યુલર રેડોક્સ્ટેસિસનું નિયમન કરવાનો આ એક માર્ગ છે, કારણ કે TAK1-ની ઉણપ NRF2 પ્રોટીન સ્તરમાં ઘટાડા સાથે સમાંતર રીતે વિવિધ માઉસ પેશીઓમાં કોઈપણ બાહ્ય ઓક્સિડન્ટની ગેરહાજરીમાં ROS ને નિયંત્રિત કરે છે [105 ].
UBA ડોમેનનો અભાવ ધરાવતી p62 રચના હજુ પણ KEAP1 ને બાંધવા માટે સક્ષમ હતી, જે સૂચવે છે કે ક્રિયાપ્રતિક્રિયા સર્વવ્યાપક KEAP1 [101] પર આધારિત નથી. જો કે, ડ્રોસોફિલા મેલાનોગાસ્ટરમાં p62 હોમોલોગ, Ref(2) નામનું, KIR મોટિફ ધરાવતું નથી અને તે DmKEAP1 સાથે સીધો સંપર્ક કરતું નથી, જો કે તે UBA ડોમેન દ્વારા સર્વવ્યાપક DmKEAP1 સાથે જોડાઈ શકે છે. વધુમાં, DmKEAP1 એટીજી 8 (સસ્તન LC3 માટે હોમોલોગ) સાથે સીધો સંપર્ક કરી શકે છે. KEAP1 ની ઉણપ એટીજી8 અને ઓટોફેજી ઇન્ડક્શનમાં પરિણમે છે જે NRF2 ઓર્થોલોગ CncC પર આધારિત છે અને TFEB/MITF [106] પર સ્વતંત્ર છે. NRF2 અને ઓટોફેજી વચ્ચેનો સંબંધ તેની કાર્યાત્મક સુસંગતતા પર પ્રકાશ પાડતા સચવાયેલો જણાય છે.
p2 દ્વારા NRF62 નું ઇન્ડક્શન KEAP1 ને બાંધવાની સ્પર્ધા અને લાઇસોસોમમાં KEAP1 ના અધોગતિનું પરિણામ છે. siRNA સાથે p62 ને સાઇલન્સ કરવાથી NRF1 અને તેના લક્ષ્ય જનીનો [2] માં ઘટાડા સાથે સમાંતર KEAP101 અર્ધ-જીવન બમણું થાય છે. કરારમાં, p62 અભિવ્યક્તિને નાબૂદ કરવાથી જંગલી પ્રકારના ઉંદરોની તુલનામાં KEAP1 ના સ્તરમાં વધારો થયો છે. ખૂબ જ સુસંગત, KEAP1 સ્તરોમાં વધારો પ્રોટીઝોમ અવરોધકો દ્વારા પ્રભાવિત થયો ન હતો પરંતુ ભૂખમરો-પ્રેરિત ઓટોફેજી [107] હેઠળ ઘટાડો થયો હતો. હકીકતમાં, KEAP1 સસ્તન પ્રાણીઓના કોષોમાં p62 અને LC3 [99], [100], [103]થી સુશોભિત ઓટોફેજિક વેસિકલ્સમાં હાજર છે. આ તમામ ડેટા સૂચવે છે કે KEAP1 એ મેક્રોઓટોફેજી મશીનરીનો સબસ્ટ્રેટ છે, પરંતુ કેટલાક વિવાદાસ્પદ પરિણામોના અસ્તિત્વને કારણે આ મુદ્દાનું વધુ વિગત સાથે વિશ્લેષણ કરવું જોઈએ. એટીજી 1-નલ ઉંદરમાં KEAP7 પ્રોટીનનું સ્તર વધ્યું હતું, જે મેક્રોઓટોફેજી [107] ના મુખ્ય અસરકર્તા છે, પરંતુ ટોરીન1, E64/પેપસ્ટાટિન અથવા બેફિલોમિસિન સાથે મેક્રોઓટોફેજીનું ફાર્માકોલોજિકલ અવરોધ KEAP1 [107], [100] એકઠા કરવામાં નિષ્ફળ ગયું હતું. એકંદરે, આ પરિણામો સૂચવે છે કે વધેલા p62 સ્તરો KEAP1 ને ઓટોફેજિક વેક્યુલોમાં અલગ કરે છે અને કદાચ આ પરિણામો KEAP1 ઓટોફેજિક ડિગ્રેડેશનમાં પરિણમે છે જે NRF2 સક્રિયકરણને મંજૂરી આપે છે (ફિગ. 3). બે અલગ-અલગ અભ્યાસોએ અહેવાલ આપ્યો છે કે આ સંદર્ભમાં સલ્ફિનીક એસિડ રિડક્ટેસિસ સેસ્ટ્રીન્સ મહત્વની ભૂમિકા ભજવે છે. SESTRIN 2 p62, KEAP1 અને RBX1 સાથે ક્રિયાપ્રતિક્રિયા કરે છે અને KEAP62 ના p1-આશ્રિત અધોગતિ અને લક્ષ્ય જનીનો NRF2 સક્રિયકરણની સુવિધા આપે છે [108]. અન્ય અભ્યાસ દર્શાવે છે કે SESTRIN 2 એ ULK1 અને p62 સાથે ક્રિયાપ્રતિક્રિયા કરી, Ser62 પર p403 ના ફોસ્ફોરાયલેશનને પ્રોત્સાહન આપ્યું જેણે KEAP1 [109] સહિત કાર્ગો પ્રોટીનના અધોગતિને સરળ બનાવ્યું.
આકૃતિ 3 NRF2 સ્તરો એડેપ્ટર પ્રોટીન p62 દ્વારા નિયંત્રિત થાય છે. mTORC351, TAK62 અથવા અન્ય કિનાસેસ દ્વારા p349 (354-DPSTGE-1) ના KIR મોટિફમાં Ser 1 નું ફોસ્ફોરાયલેશન NRF1 માં ETGE મોટિફ સાથે સામ્યતાના કારણે KEAP2 સાથે બંધન માટે વધુ આકર્ષણમાં પરિણમે છે. પરિણામે, ફોસ્ફોરીલેટેડ p62 NRF2 ને વિસ્થાપિત કરે છે અને KEAP1 ને બાંધે છે. p62 માં LIR મોટિફ ઓટોફેગોસોમલ મેમ્બ્રેનમાં LC3 સાથે ક્રિયાપ્રતિક્રિયાને સક્ષમ કરે છે, જેથી p62-KEAP1 સંકુલ આખરે લાઇસોસોમમાં અધોગતિ પામે છે. પરિણામે NRF2 એ એકઠા કરવામાં, ન્યુક્લિયસમાં સ્થાનાંતરિત કરવામાં અને P62 સહિત ARE-સમાવતી જનીનોના ટ્રાન્સક્રિપ્શનને વધારવામાં સક્ષમ છે. આ નિયમનકારી મિકેનિઝમ ટકાઉ NRF2 પ્રતિભાવ પ્રદાન કરે છે, કારણ કે NRF1 પ્રવૃત્તિને અટકાવવા માટે KEAP2 ને નવા સંશ્લેષણની જરૂર છે.
NRF2 દ્વારા મેક્રોઓટોફેજી જીન્સનું મોડ્યુલેશન
NRF2 મેક્રોઓટોફેજી માટે સંબંધિત જનીનોની અભિવ્યક્તિનું નિયમન કરે છે તેમજ તે UPR અને UPS માટે કરે છે. પ્રથમ પુરાવા એવા અભ્યાસોમાંથી આવ્યા છે જેમાં p62 અભિવ્યક્તિ ઇલેક્ટ્રોફાઇલ્સ, ROS અને નાઈટ્રિક ઓક્સાઇડ [110], [111], [112]ના સંપર્કમાં આવવાથી પ્રેરિત હોવાનું દર્શાવવામાં આવ્યું હતું. ઇન્ડક્શનની મિકેનિઝમનું વર્ણન કેટલાક વર્ષો પછી એ શોધ સાથે કરવામાં આવ્યું હતું કે p62 તેના જનીન પ્રમોટર [99] માં કાર્યાત્મક ARE ધરાવે છે. તાજેતરના અભ્યાસમાં, બાયોઇન્ફોર્મેટિક્સ પૃથ્થકરણ અને ચિપ એસેસને પગલે અન્ય કેટલાંક કાર્યાત્મક ARE મળી આવ્યા અને માન્ય કરવામાં આવ્યા. વધુમાં, Nrf2-નોકઆઉટ ઉંદરના માઉસ એમ્બ્રીયોનિક ફાઈબ્રોબ્લાસ્ટ્સ અને કોર્ટિકલ ન્યુરોન્સમાં ઘટાડો p62 અભિવ્યક્તિ દર્શાવવામાં આવી હતી, જેને NRF2-વ્યક્ત લેન્ટીવાયરસથી બચાવી શકાય છે. એ જ રીતે, NRF2 ની ઉણપએ ઉંદર હિપ્પોકેમ્પસ [62] થી ઇજાગ્રસ્ત ચેતાકોષોમાં p36 સ્તરમાં ઘટાડો કર્યો. તેથી, એવું સૂચવવામાં આવ્યું છે કે NRF2 સક્રિયકરણ p62 સ્તરમાં વધારો કરે છે, પરિણામે KEAP1 અધોગતિ થાય છે અને હકારાત્મક પ્રતિસાદ લૂપમાં વધુ NRF2 સ્થિરીકરણની તરફેણ કરે છે. NRF2 ઇન્ડક્શનની આ બિન-પ્રમાણિક પદ્ધતિને જનીન અભિવ્યક્તિમાં ફેરફારની જરૂર છે અને તે લાંબા સમય સુધી સેલ્યુલર તણાવ માટે સંબંધિત પ્રતિભાવ હોઈ શકે છે.
કાર્ગો રેકગ્નિશન પ્રોટીન NDP52 ને NRF2 દ્વારા ટ્રાન્સક્રિપ્શનલી રેગ્યુલેટેડ બતાવવામાં આવ્યું હતું. NDP52 p62 ની સમાન રીતે કામ કરે છે, સર્વવ્યાપક પ્રોટીનને ઓળખે છે અને LIR ડોમેન દ્વારા LC3 સાથે ક્રિયાપ્રતિક્રિયા કરે છે, જેથી લાઇસોસોમ્સમાં કાર્ગો ડિગ્રેડ થાય છે. Ndp52 પ્રમોટર DNA ક્રમમાં પાંચ પુટેટિવ ARE મળી આવ્યા હતા. તેમાંથી ત્રણને NRF2-મધ્યસ્થી Ndp52 ટ્રાંસ્ક્રિપ્શન [113] માટે અનિવાર્ય તરીકે અલગ-અલગ મ્યુટન્ટ રચનાઓ અને ChIP એસેઝ સાથે ઓળખવામાં આવ્યા હતા. નોંધનીય છે કે, Nrf52-નોકઆઉટ ઉંદરના હિપ્પોકેમ્પસમાં Ndp2 mRNA સ્તરમાં ઘટાડો થયો હતો. NRF2-નિયંત્રિત ARE [36] તરીકે સ્વતંત્ર અભ્યાસમાં આમાંથી એક ક્રમ પણ માન્ય કરવામાં આવ્યો હતો.
જો કે, ઓટોફેજીના મોડ્યુલેશનમાં NRF2 ની ભૂમિકા આ બે કાર્ગો-ઓળખાણ પ્રોટીનના ઇન્ડક્શન સુધી મર્યાદિત નથી. વધારાના ઓટોફેજી-સંબંધિત જનીનોના મોડ્યુલેશનમાં NRF2 ની ભૂમિકામાં ઊંડી સમજ મેળવવા માટે, અમારા જૂથે બે પ્રોટીન, MAFK અને BACH1 માટે ક્રોમેટિન ઇમ્યુનોપ્રિસિપિટેશન ડેટાબેઝ ENCODE નું સ્ક્રીનીંગ કર્યું, જે NRF2-નિયમિત ARE ને બાંધે છે. JASPAR ની સર્વસંમતિ ARE ક્રમમાંથી જનરેટ થયેલ સ્ક્રિપ્ટનો ઉપયોગ કરીને, અમે ઘણા ઓટોફેજી જનીનોમાં કેટલાક પુટેટિવ ARE ને ઓળખ્યા. આમાંથી બાર સિક્વન્સને નવ ઓટોફેજી જનીનોમાં NRF2 રેગ્યુલેટેડ ARE તરીકે માન્ય કરવામાં આવ્યા હતા, જેની અભિવ્યક્તિ Nrf2-નોકઆઉટ ઉંદરના માઉસ એમ્બ્રીયો ફાઈબ્રોબ્લાસ્ટ્સમાં ઓછી થઈ હતી પરંતુ NRF2- વ્યક્ત કરનાર લેન્ટીવાયરસ દ્વારા પુનઃસ્થાપિત કરી શકાય છે. અમારા અભ્યાસે દર્શાવ્યું છે કે NRF2 ઓટોફેજી ઇનિશિયેશન (ULK1), કાર્ગો રેકગ્નિશન (p62 અને NDP52), ઓટોફેગોસોમ ફોર્મેશન (ATG4D, ATG7 અને GABARAPL1), વિસ્તરણ (ATG2B અને ATG5B સહિત ઓટોફેજીક પ્રક્રિયાના વિવિધ તબક્કામાં સંકળાયેલા કેટલાક જનીનોની અભિવ્યક્તિને સક્રિય કરે છે. ), અને ઓટોલિસોસોમ ક્લિયરન્સ (ATG4D). પરિણામે, જ્યારે NRF2 ગેરહાજર હતું ત્યારે હાઇડ્રોજન પેરોક્સાઇડના પ્રતિભાવમાં ઓટોફેજી પ્રવાહ ક્ષતિગ્રસ્ત હતો [36].
ખામીયુક્ત ઓટોફેજી અનેક ન્યુરોડીજનરેટિવ રોગોમાં મહત્વની ભૂમિકા ભજવતી હોવાનું દર્શાવવામાં આવ્યું છે [114] અને ઓટોફેજીને નાબૂદ કરવાથી ઉંદર [115], [116] માં ન્યુરોડિજનરેશન તરફ દોરી જાય છે. એટીજી7-નોકઆઉટ ઉંદરે જાહેર કર્યું કે ઓટોફેજીની ઉણપ ubiquitin-પોઝિટિવ ઇન્ક્લુઝન બોડીમાં p62 સંચયમાં પરિણમે છે. KEAP1 ને આ સમાવિષ્ટ સંસ્થાઓમાં અલગ કરવામાં આવ્યું હતું, જે NRF2 સ્થિરીકરણ તરફ દોરી જાય છે અને લક્ષ્ય જનીનોને ઇન્ડક્શન કરે છે [103]. અગત્યની રીતે, સર્વવ્યાપક પ્રોટીન સાથે p62 નું અતિશય સંચય AD, PD અને ALS [117] સહિત ન્યુરોડિજનરેટિવ રોગોમાં ઓળખવામાં આવ્યું છે. વાસ્તવમાં, એડી દર્દીઓના ઉચ્ચ સ્તરના એપીપી અથવા ટીએયુને વ્યક્ત કરતા ચેતાકોષોએ પણ p62 અને ન્યુક્લિયર NRF2 વ્યક્ત કર્યા હતા, જે ઓટોફેજી [36] દ્વારા ઇન્ટ્રાન્યુરોનલ એગ્રીગેટ્સને ડિગ્રેજ કરવાના તેમના પ્રયાસનું સૂચન કરે છે.
NRF2 ની ઉણપ AD ના સંદર્ભમાં પ્રોટીન એકત્રીકરણને વધારે છે. હકીકતમાં, ફોસ્ફોરીલેટેડ અને સાર્કોસિલ-અદ્રાવ્ય TAU ના વધેલા સ્તરો Nrf2-નોકઆઉટ ઉંદરમાં જોવા મળે છે, જો કે જંગલી-પ્રકારની પૃષ્ઠભૂમિ [113] સાથે સરખામણી કરતાં કિનેઝ અથવા ફોસ્ફેટેઝ પ્રવૃત્તિઓમાં કોઈ તફાવત શોધી શકાયો નથી. મહત્ત્વપૂર્ણ રીતે, NDP52 એ મ્યુરિન ચેતાકોષોમાં TAU સાથે સહ-સ્થાનિકીકરણ માટે દર્શાવવામાં આવ્યું હતું અને ફોસ્ફો-TAU અને NDP52 વચ્ચેની સીધી ક્રિયાપ્રતિક્રિયા ઉંદર અને AD બંને નમૂનાઓમાં સહ-રોગપ્રતિકારક પ્રયોગો દ્વારા દર્શાવવામાં આવી હતી, TAU અધોગતિમાં તેની ભૂમિકા તરફ નિર્દેશ કરે છે. રસપ્રદ વાત એ છે કે, ચેતાકોષોમાં NDP52, p62 અથવા NRF2 ને શાંત પાડવાથી ફોસ્ફો-ટીએયુ [113], [118] માં વધારો થયો છે. વધુમાં, જ્યારે NRF1 ગેરહાજર હતું ત્યારે APP/PS9?E2 ઉંદરના હિપ્પોકેમ્પસમાં વધેલા ઇન્ટ્રાન્યુરોનલ એપીપી એગ્રીગેટ્સ જોવા મળ્યા હતા. આ બદલાયેલા ઓટોફેજી માર્કર્સ સાથે સહસંબંધ ધરાવે છે, જેમાં ફોસ્ફો-mTOR/mTOR અને ફોસ્ફો-p70S6k/p70S6k ગુણોત્તર (ઓટોફેજી નિષેધનું સૂચક), પ્રી-કેથેપ્સિન ડીના વધેલા સ્તરો અને મોટી સંખ્યામાં મલ્ટિવેસિક્યુલર બોડીઝ [119]નો સમાવેશ થાય છે. માનવ APP (V717I) અને TAU (P301L) સહ-અભિવ્યક્ત કરતા ઉંદરમાં, NRF2 ની ઉણપને કારણે અદ્રાવ્ય અપૂર્ણાંકમાં કુલ અને ફોસ્ફો-ટીએયુના સ્તરમાં વધારો થયો અને p62, NDP52, ULK1, ULK5, ના ન્યુરોનલ સ્તરોમાં ઘટાડો સાથે ઇન્ટ્રાન્યુરોનલ એપીપી એગ્રીગેટ્સમાં વધારો થયો. ATG1 અને GABARAPL62. એડેપ્ટર પ્રોટીન p2 અને APP અથવા TAU વચ્ચેનું સહ-સ્થાનિકીકરણ NRF36 [2] ની ગેરહાજરીમાં ઘટાડવામાં આવ્યું હતું. એકંદરે, આ પરિણામો ન્યુરોનલ ઓટોફેજીમાં NRFXNUMX નું મહત્વ દર્શાવે છે.
પ્રોટીઓસ્ટેસિસને મોડ્યુલેટ કરવા માટે વિવિધ ટ્રાન્સક્રિપ્શન પરિબળો સંકલનથી કાર્ય કરે છે
સ્થિર સ્થિતિની સ્થિતિમાં, પ્રોટીઓસ્ટેસિસ પ્રોટીન-પ્રોટીન ક્રિયાપ્રતિક્રિયાઓ અને ઝડપી પ્રતિસાદ પ્રાપ્ત કરીને અનુવાદ પછીના ફેરફારો દ્વારા નિયંત્રિત થાય છે. જો કે, સેલ્યુલર અનુકૂલન માટે યુપીઆર, યુપીએસ અને ઓટોફેજી જનીનોના ટ્રાન્સક્રિપ્શનલ નિયમનની જરૂર છે. ચેતા કોષો ઓક્સિડેટીવ અને પ્રોટીટોક્સિક તણાવ સહિત નીચા-ગ્રેડના ઝેરી અપમાનને સતત સબમિટ કરે છે તે ધ્યાનમાં લેતા, ટ્રાન્સક્રિપ્શનલ મોડ્યુલેશન દ્વારા પ્રેરિત પ્રોટીઓસ્ટેસિસનું મજબૂતીકરણ મગજના અધોગતિને રોકવામાં મદદ કરી શકે છે.
યુપીઆરના કિસ્સામાં, ત્રણેય હાથમાંથી દરેકનું સક્રિયકરણ આખરે ચોક્કસ જનીનોના ટ્રાન્સક્રિપ્શનલ ઇન્ડક્શનમાં પરિણમશે ([43] માં સમીક્ષા). દાખલા તરીકે, ATF6-ઉત્પન્ન ફ્રેગમેન્ટ (ATF6f) ER-સ્ટ્રેસ રિસ્પોન્સ એલિમેન્ટ્સ (ERSE) સાથે જોડાય છે અને XBPI, BIP અને CHOP સહિત અનેક જનીનોની અભિવ્યક્તિને પ્રેરિત કરે છે. વધુમાં, PERK સિગ્નલિંગ ટ્રાન્સક્રિપ્શન ફેક્ટર ATF4 ના સક્રિયકરણ તરફ દોરી જાય છે, જે બહુવિધ UPR-સંબંધિત જનીનો અને NRF2 લક્ષ્ય જનીનો Hmox1 અને p62 સહિત કેટલાક અન્યની અભિવ્યક્તિને નિયંત્રિત કરે છે. છેલ્લે, IRE1 સક્રિયકરણ સક્રિય ટ્રાન્સક્રિપ્શન પરિબળ, સ્પ્લિસ્ડ XBP1 (XBP1s) ની રચનામાં પરિણમે છે, જે પ્રોટીન ફોલ્ડિંગમાં સામેલ જીન્સ એન્કોડિંગ પ્રોટીનના ટ્રાન્સક્રિપ્શનને નિયંત્રિત કરે છે.
બીજી બાજુ, NRF1 મગજમાં પ્રોટીસોમલ જનીન અભિવ્યક્તિ માટે જરૂરી હોવાનું દર્શાવવામાં આવ્યું હતું, કારણ કે Nrf1-નોકઆઉટ ઉંદર 20S કોરના વિવિધ સબ્યુનિટ્સને એન્કોડ કરતા જનીનોની ઘટેલી અભિવ્યક્તિ દર્શાવે છે, તેમજ 19S રેગ્યુલેટરી કોમ્પ્લેક્સ સાથે ક્ષતિગ્રસ્ત પ્રોટીસોમલ કાર્ય [90] ]. NRF1 અને NRF2 બંને તેના લક્ષ્ય જનીનોના પ્રમોટર પ્રદેશોમાં ARE સિક્વન્સ સાથે જોડાય છે, જે સૂચવે છે કે તેઓ ટ્રાન્સક્રિપ્શનલ પ્રવૃત્તિઓને ઓવરલેપ કરે છે, જો કે તેઓ તેમની નિયમનકારી પદ્ધતિઓ અને સેલ્યુલર સ્થાનિકીકરણ [120] માં અલગ છે.
ફોર્કહેડ બોક્સ O (FOXO) પરિવારના ટ્રાન્સક્રિપ્શન પરિબળો બહુવિધ ઓટોફેજી-સંબંધિત જનીનોની અભિવ્યક્તિને નિયંત્રિત કરે છે. NRF2 સાથે જે થાય છે તેની જેમ, FOXO સભ્યોની પ્રવૃત્તિના નિયમનના બહુવિધ સ્તરો છે, જે પોષક અથવા ઓક્સિડેટીવ તણાવ [121] પર પ્રેરિત થઈ શકે છે. છેલ્લે, ટ્રાંસ્ક્રિપ્શન પરિબળ TFEB, જે લાઇસોસોમલ બાયોજેનેસિસના મુખ્ય નિયમનકાર તરીકે ગણવામાં આવે છે, તે પોષક તાણની પરિસ્થિતિઓ હેઠળ ઓટોફેજીના નિયમનમાં નિર્ણાયક ભૂમિકા ભજવે છે. આમ, mTORC1 નું નિષેધ TFEB ના પરમાણુ સ્થાનાંતરણ તરફ દોરી જાય છે અને ઓટોફેજી જીન્સ [122] ની અભિવ્યક્તિ ઇન્ડક્શન તરફ દોરી જાય છે.
એકંદરે, આ મશીનરીઓના વિવિધ ટ્રાન્સક્રિપ્શનલ રેગ્યુલેટરનું અસ્તિત્વ પણ ક્રોસસ્ટૉક અને અંશતઃ રીડન્ડન્ટ મિકેનિઝમ્સ સૂચવે છે જે વિવિધ સંજોગોમાં પ્રોટીઓસ્ટેસિસની ખાતરી આપી શકે છે. તદનુસાર, NRF2 ની પેશીઓમાં સંબંધિત ભૂમિકા હોઈ શકે છે જે ઓક્સિડેટીવ તાણના ઉચ્ચ સ્તરને સમર્થન આપે છે. દાખલા તરીકે, ઓક્સિડેટીવ સ્ટ્રેસ-પ્રેરિત NRF2 પોષક તત્ત્વોથી ભરપૂર પરિસ્થિતિઓમાં કામ કરી શકે છે, જે ઓટોફેજીને ટ્રાન્સક્રિપ્શનલી અપ-રેગ્યુલેટ કરે છે, જે ભૂખમરાની સ્થિતિમાં TFEB માટે જોવા મળ્યું છે. તદુપરાંત, મગજ મોટાભાગે પોષક તત્ત્વોથી ભરપૂર પરિસ્થિતિઓ હેઠળ કાર્ય કરે છે, જે NRF2 ને ન્યુરોન્સમાં ઓટોફેજીને સક્રિય કરવા માટે સંબંધિત પદ્ધતિ તરીકે રજૂ કરે છે.
પ્રોટીનોપેથીમાં NRF2 માટે આશાસ્પદ ઉપચારાત્મક સંભવિત
છેલ્લાં કેટલાંક વર્ષોમાં, NRF2 પ્રવૃત્તિ પર UPR, UPS અને ઑટોફેજીની નિયમનકારી ભૂમિકાઓ તેમજ આ ત્રણ પ્રણાલીઓના ઘટકોના પારસ્પરિક NRF2-મધ્યસ્થી ટ્રાન્સક્રિપ્શનના જ્ઞાનમાં મોટી પ્રગતિ થઈ છે. તેથી, ન્યુરોડિજનરેટિવ રોગોમાં પ્રોટીન ક્લિયરન્સના નિર્ણાયક નિયમનકાર તરીકે NRF2 ના શોષણના આધારે નવી ઉપચારાત્મક શક્યતાઓ ઊભી થઈ શકે છે.
જો કે, બાકીનો એક મુખ્ય પ્રશ્ન એ છે કે મગજમાં NRF2 સ્તર વધારવા માટે તે ઉપયોગી છે કે નુકસાનકારક છે. રોગચાળા સંબંધી માહિતીનું વિશ્લેષણ આંશિક જવાબ આપી શકે છે, કારણ કે તે સૂચવે છે કે NFE2L2 જનીન અત્યંત પોલીમોર્ફિક છે અને તેના પ્રમોટર રેગ્યુલેટરી પ્રદેશમાં જોવા મળતા કેટલાક એકલ ન્યુક્લિયોટાઈડ પોલીમોર્ફિઝમ્સ વસ્તીના સ્તરે અને કેટલાક હેપ્લોટાઈપ્સ અને કેટલાક હેપ્લોટાઈપ્સ જનીન અભિવ્યક્તિમાં વિવિધતા પ્રદાન કરી શકે છે. AD, PD અથવા ALS [123] ના જોખમમાં ઘટાડો અને/અથવા વિલંબિત શરૂઆત સાથે સંકળાયેલા હતા. વધુમાં, હેયસ અને સહકર્મીઓ [124] દ્વારા ચર્ચા કર્યા મુજબ, NRF2 અસરમાં U-આકારનો પ્રતિભાવ હોઈ શકે છે, એટલે કે ખૂબ નીચા NRF2 સ્તરને કારણે સાયટોપ્રોટેક્શનની ખોટ થઈ શકે છે અને સ્ટ્રેસર્સ પ્રત્યે સંવેદનશીલતા વધી શકે છે, જ્યારે વધુ પડતું NRF2 હોમિયોસ્ટેટિક સંતુલનને ખલેલ પહોંચાડી શકે છે. રિડક્ટિવ સિનેરીયો (રિડક્ટિવ સ્ટ્રેસ), જે પ્રોટીન મિસફોલ્ડિંગ અને એકત્રીકરણની તરફેણ કરશે. મગજમાં નીચા NRF2 સ્તરો એ વિચારને સમર્થન આપે છે કે પેથોલોજીકલ પરિસ્થિતિઓમાં લાભ મેળવવા માટે થોડું અપ-નિયમન પૂરતું હોઈ શકે છે. વાસ્તવમાં, પ્રોટીન ક્લિયરન્સના ફાર્માકોલોજિકલ NRF2-મધ્યસ્થી સક્રિયકરણની રક્ષણાત્મક ભૂમિકા વિવિધ ન્યુરોડિજનરેશન સેલ કલ્ચર અને વિવો મોડલ્સમાં દર્શાવવામાં આવી છે.
SFN એ ફાર્માકોલોજિકલ NRF2 એક્ટિવેટર છે જે પ્રોટીસોમલ અને ઓટોફેજી જનીન અભિવ્યક્તિ [95], [36] પ્રેરિત કરવા માટે દર્શાવવામાં આવ્યું હતું. રસપ્રદ રીતે, જો અને સહકર્મીઓએ દર્શાવ્યું કે SFN એ ફોસ્ફોરીલેટેડ TAU નું સ્તર ઘટાડ્યું અને Beclin-1 અને LC3-II માં વધારો કર્યો, સૂચવે છે કે NRF2 સક્રિયકરણ ઓટોફેજી [113] દ્વારા આ ઝેરી પ્રોટીનના અધોગતિને સરળ બનાવી શકે છે. વધુમાં, MHtt ના અધોગતિને SFN સાથે ઉન્નત કરવામાં આવ્યું હતું, અને આ MG132 ના ઉપયોગથી પાછું લાવવામાં આવ્યું હતું, જે આ ઝેરી પ્રોટીન [95] ના પ્રોટીસોમલ અધોગતિ દર્શાવે છે. ફોસ્ફો- અને અદ્રાવ્ય-ટીએયુનું ઓટોફેજી-મધ્યસ્થી અધોગતિ ઓર્ગેનિક ફ્લેવોનોઈડ ફિસેટિન સાથે નોંધવામાં આવી હતી. આ સંયોજન તેના કેટલાક લક્ષ્ય જનીનો સાથે TFEB અને NRF2 બંનેના સક્રિયકરણ અને પરમાણુ સ્થાનાંતરણને વારાફરતી પ્રોત્સાહન આપીને ઓટોફેજીને પ્રેરિત કરવામાં સક્ષમ હતું. આ પ્રતિભાવને TFEB અથવા NRF2 મૌન [125] દ્વારા અટકાવવામાં આવ્યો હતો. બોટ અને સહકર્મીઓએ કરોડરજ્જુ અને બલ્બર મસ્ક્યુલર એટ્રોફીમાં પ્રોટીનની ઝેરી અસર પર એક સાથે NRF2, NRF1 અને HSF1 એક્ટિવેટરની ફાયદાકારક અસરોની જાણ કરી, જે પોલીગ્લુટામાઇન-એન્કોડિંગ CAG પુનરાવર્તનના વિસ્તરણને કારણે ન્યુરોડિજનરેટિવ ડિસઓર્ડર છે જેમાં પ્રોટીન એગ્રીગેટ્સ હાજર છે [126]. મલ્ટિપલ સ્ક્લેરોસિસ [2], [12] ની સારવાર માટે NRF2 ઇન્ડ્યુસર ડાયમિથાઈલ ફ્યુમરેટ (DMF) ની મૌખિક રચના, BG-127 ની મંજૂરી સાથે ન્યુરોડિજનરેટિવ ડિસઓર્ડરની સારવાર માટે NRF128 સક્રિયકરણની સંભવિતતા દર્શાવવામાં આવી છે. મજબૂત બળતરા ઘટક સાથે સ્વયંપ્રતિરક્ષા બિમારીઓ સાથે ડીએમએફની સફળતા સૂચવે છે કે ન્યુરોડિજનરેટિવ રોગો આ દવાને ફરીથી સ્થાનાંતરિત કરવાથી ફાયદો થઈ શકે છે. PD ના ?-synucleinopathy મોડલના તાજેતરના પ્રીક્લિનિકલ અભ્યાસમાં, DMF ને ઓટોફેજી [129] ના ઇન્ડક્શનને કારણે ન્યુરોપ્રોટેક્ટીવ હોવાનું દર્શાવવામાં આવ્યું હતું. ન્યુરોડિજનરેશન પર NRF2 ની ફાયદાકારક અસરોની જાણ કરતા અભ્યાસો પરંતુ પ્રોટીન ક્લિયરન્સ પર તેની અસર પર ધ્યાન કેન્દ્રિત કરતા નથી તે વધુ વિપુલ પ્રમાણમાં છે (વ્યાપક સમીક્ષા માટે, જુઓ [7]). આ તદ્દન સુસંગત છે, કારણ કે તે બહુવિધ નુકસાનકારક પ્રક્રિયાઓને હાઇલાઇટ કરે છે જેને NRF2 માં એક જ હિટ દ્વારા એકસાથે લક્ષ્ય બનાવી શકાય છે, જેમાં ઓક્સિડેટીવ તણાવ, ન્યુરોઇન્ફ્લેમેશન અથવા મિટોકોન્ડ્રીયલ ડિસફંક્શનનો પણ સમાવેશ થાય છે. જો કે, મગજમાં ઝેરી પ્રોટીનના અધોગતિને સરળ બનાવવા માટે NRF2 નું ફાર્માકોલોજિકલ સક્રિયકરણ એક માન્ય વ્યૂહરચના હોઈ શકે છે કે કેમ તે નિશ્ચિતપણે નક્કી કરવા માટે ભવિષ્યના કાર્યની જરૂર પડશે.
અગાઉ સમજાવ્યા મુજબ, GSK-3 વધી ગયું? ન્યુરોડિજનરેટિવ રોગોમાં પ્રવૃત્તિની જાણ કરવામાં આવી હતી અને એવું અનુમાન કરવામાં આવ્યું છે કે પરિણામી NRF2 ઘટાડો નુકસાનકારક પરિણામ માટે આંશિક રીતે જવાબદાર હોઈ શકે છે. આ રોગવિજ્ઞાનવિષયક પરિસ્થિતિઓ હેઠળ, GSK-3 અવરોધકો પણ NRF2 સ્તર અને પ્રોટીઓસ્ટેસિસ વધારવા માટે સહકાર આપી શકે છે. જીએસકે-3 અવરોધકોની ફાયદાકારક અસરો ન્યુરોડીજનરેશનના વિવિધ મોડલમાં નોંધવામાં આવી છે અને વધુ રસપ્રદ, જીએસકે-3 દમન ઝેરી પ્રોટીન [130], [131], [132], [133]ના સ્તરને ઘટાડવા માટે દર્શાવવામાં આવ્યું હતું. જોકે GSK-3 અવરોધ અને NRF2-પ્રોટીઓસ્ટેસિસને પ્રોત્સાહન આપતા જનીનોના ટ્રાન્સક્રિપ્શનલ નિયમન વચ્ચે કોઈ સીધો સંબંધ હજુ સુધી જોવા મળ્યો નથી, એવું અનુમાન કરવું વાજબી છે કે GSK-3 પ્રવૃત્તિના ડાઉન-રેગ્યુલેશનના પરિણામે NRF2 સ્તરમાં વધારો થશે, જે આખરે પ્રબલિત થશે. પ્રોટીઓસ્ટેસિસ
NRF2 ની ટ્રાન્સક્રિપ્શનલ પ્રવૃત્તિ તેમજ પ્રોટીઓસ્ટેસિસ જાળવવાની સેલ્યુલર ક્ષમતા વય સાથે ઘટે છે, જે ન્યુરોડિજનરેટિવ રોગોના વિકાસ માટેનું મુખ્ય જોખમ પરિબળ છે. તે વિચારવું વાજબી છે કે NRF2 નું મજબૂતીકરણ અને પરિણામે, પ્રોટીઓસ્ટેસિસ, ઓછામાં ઓછું, પ્રોટીન એકત્રીકરણ અને ન્યુરોડિજનરેશનના સંચયમાં વિલંબ કરશે. ખરેખર, 18?-glycyrrhetinic એસિડ (18?-GA) ટ્રાઇટરપેનોઇડ સાથે માનવ સેન્સેન્ટ ફાઇબ્રોબ્લાસ્ટની સારવારથી NRF2 સક્રિયકરણને પ્રોત્સાહન મળે છે, જે પ્રોટીઝોમ ઇન્ડક્શન તરફ દોરી જાય છે અને આયુષ્યમાં વધારો કરે છે. આ અભ્યાસ સૂચવે છે કે NRF2 નું ફાર્માકોલોજીકલ સક્રિયકરણ જીવનના અંતમાં પણ શક્ય છે [86]. તદુપરાંત, પછીના અભ્યાસે દર્શાવ્યું હતું કે આ સંયોજન SKN-1 અને C.elegans માં પ્રોટીઝોમ સક્રિયકરણની મધ્યસ્થી કરે છે અને સંબંધિત નેમાટોડ મોડલ્સ [134] માં AD ની પ્રગતિ પર ફાયદાકારક અસર કરે છે.
તમામ બાબતોને ધ્યાનમાં લેવામાં આવે છે, પ્રોટીઓસ્ટેસિસ-સંબંધિત જનીનોનું NRF2- મધ્યસ્થી ઇન્ડક્શન વિવિધ પ્રોટીનોપેથીઓમાં ફાયદાકારક જણાય છે.
સલ્ફોરાફેન અને કેન્સર, મૃત્યુદર, વૃદ્ધત્વ, મગજ અને વર્તન, હૃદય રોગ અને વધુ પર તેની અસરો
Isothiocyanates એ કેટલાક સૌથી મહત્વપૂર્ણ છોડ સંયોજનો છે જે તમે તમારા આહારમાં મેળવી શકો છો. આ વિડિયોમાં હું તેમના માટે અત્યાર સુધીનો સૌથી વ્યાપક કેસ બનાવું છું. ટૂંકા ધ્યાન ગાળો? નીચેના ટાઈમ પોઈન્ટ્સમાંથી કોઈ એક પર ક્લિક કરીને તમારા મનપસંદ વિષય પર જાઓ. નીચે સંપૂર્ણ સમયરેખા.
મુખ્ય વિભાગો:
00:01:14 - કેન્સર અને મૃત્યુદર
00:19:04 - વૃદ્ધત્વ
00:26:30 - મગજ અને વર્તન
00:38:06 - અંતિમ રીકેપ
00:40:27 - માત્રા
સંપૂર્ણ સમયરેખા:
00:00:34 - સલ્ફોરાફેનનો પરિચય, વિડીયોનું મુખ્ય ધ્યાન.
00:01:14 - ક્રુસિફેરસ શાકભાજીનો વપરાશ અને સર્વ-કારણ મૃત્યુદરમાં ઘટાડો.
00:44:24 - શાકભાજીમાંથી સક્રિય માયરોસિનેઝ સાથે જોડવામાં આવે ત્યારે પૂરક વધુ સારી રીતે કામ કરે છે.
00:44:56 - રાંધવાની તકનીકો અને ક્રુસિફેરસ શાકભાજી.
00:46:06 - ગોઇટ્રોજન તરીકે આઇસોથિયોસાયનેટ્સ.
ન્યુક્લિયર ફેક્ટર એરિથ્રોઇડ-ડેરિવ્ડ 2 (NF-E2)-સંબંધિત પરિબળ 2, અન્યથા Nrf2 તરીકે ઓળખાય છે, તે એક ટ્રાન્સક્રિપ્શન પરિબળ છે જે વિવિધ એન્ટીઑકિસડન્ટ અને ડિટોક્સિફાઇંગ એન્ઝાઇમ્સની અભિવ્યક્તિને નિયંત્રિત કરે છે. સંશોધન અભ્યાસોએ ઓક્સિડેટીવ તણાવને નિયંત્રિત કરવામાં તેની ભૂમિકા પણ દર્શાવી છે. મોટાભાગના ન્યુરોડિજનરેટિવ રોગો, જેમ કે અલ્ઝાઈમર રોગ અને પાર્કિન્સન રોગ, ઓક્સિડેટીવ તણાવ અને ક્રોનિક બળતરા દ્વારા વર્ગીકૃત થયેલ છે, જેનું સામાન્ય લક્ષ્ય છે. Nrf2 સારવાર અભિગમ. ડૉ. એલેક્સ જિમેનેઝ ડીસી, CCST ઇનસાઇટ
સમાપન નોંધ, ઉપસંહાર
ટ્રાન્સક્રિપ્શન ફેક્ટર NRF2 યુપીઆર, યુપીએસ અને ઓટોફેજી (ફિગ. 4) માં ફેરફારોને સેન્સિંગ અને મોડ્યુલેટ કરીને પ્રોટીઓસ્ટેટિક પ્રતિભાવનું આયોજન કરે છે. પરિણામે, NRF2 નો અભાવ પ્રોટીનોપેથીમાં વધારો કરવા માટે દર્શાવવામાં આવ્યો છે, જે સૂચવે છે કે શ્રેષ્ઠ પ્રોટીન ક્લિયરન્સ માટે NRF2 જરૂરી છે. બધા એકસાથે, અમે અનુમાન કરી શકીએ છીએ કે NRF2 પ્રોટીનોપેથી માટે એક રસપ્રદ ઉપચારાત્મક લક્ષ્ય હોઈ શકે છે.
આકૃતિ 4 NRF2 પ્રોટીટોક્સિક-પ્રાપ્ત કટોકટી સંકેતોને રક્ષણાત્મક ટ્રાન્સક્રિપ્શનલ પ્રતિભાવ સાથે જોડતા હબ તરીકે. અનફોલ્ડ/મીસફોલ્ડ પ્રોટીનનું સંચય ER માં અનફોલ્ડ પ્રોટીન પ્રતિભાવ (UPR) ના સક્રિયકરણ તરફ દોરી જશે. PERK અથવા MAPK નું સક્રિયકરણ ER-નિવાસી Gpx8 ના ટ્રાન્સક્રિપ્શનલ ઇન્ડક્શનમાં પરિણમી શકે છે અને GSH સ્તરને નિયંત્રિત કરતા ઘણા ઉત્સેચકો, યોગ્ય પ્રોટીન ફોલ્ડિંગની ખાતરી કરવા માટે મહત્વપૂર્ણ છે. પ્રોટીન એગ્રીગેટ્સ પ્રોટીઝોમ એક્ટિવિટી (યુપીએસ) ને અટકાવે છે, સંભવતઃ NRF2 ડિગ્રેડેશનને ટાળે છે. NRF2 ખાસ કરીને Psma3, Psma6, Psmb1, Psmb5 અને પોમ્પ જનીનોના ટ્રાન્સક્રિપ્શનને મોડ્યુલેટ કરવા માટે દર્શાવવામાં આવ્યું છે. D2T ના પ્રતિભાવમાં NRF3-આશ્રિત રીતે કેટલાક અન્ય સબ્યુનિટ્સ ઉપર-નિયમિત કરવામાં આવ્યા હતા, સંભવતઃ NRF2 દ્વારા નિયમન કરાયેલ પ્રોટીઝોમ સબ્યુનિટ્સની સૂચિને વિસ્તૃત કરી હતી. પ્રોટીન એકત્રીકરણના અધોગતિ માટે ઓટોફેજી એ મુખ્ય માર્ગ છે. ઓટોફેજી NRF2 ને પણ નિયંત્રિત કરે છે, p2, Ndp62, Ulk52, Atg1b, Atg2c, Atg4, Atg5 અને Gabarapl7 ના NRF1 ટ્રાન્સક્રિપ્શનલ ઇન્ડક્શન સાથે આ ડિગ્રેડેશન પાથવેને જોડે છે.
ઉપરના લેખ મુજબ, જ્યારે ન્યુરોડિજનરેટિવ રોગોના લક્ષણોની સારવાર વિવિધ સારવાર વિકલ્પો દ્વારા કરી શકાય છે, સંશોધન અભ્યાસોએ દર્શાવ્યું છે કે Nrf2 સક્રિયકરણ મદદરૂપ સારવાર અભિગમ હોઈ શકે છે. કારણ કે Nrf2 એક્ટિવેટર્સ રોગની વ્યાપક પદ્ધતિઓને લક્ષ્ય બનાવે છે, તમામ ન્યુરોડિજનરેટિવ રોગો Nrf2 ટ્રાન્સક્રિપ્શન પરિબળના ઉપયોગથી લાભ મેળવી શકે છે. Nrf2 ના તારણોએ ન્યુરોડિજનરેટિવ રોગોની સારવારમાં ક્રાંતિ લાવી છે. અમારી માહિતીનો અવકાશ ચિરોપ્રેક્ટિક અને કરોડરજ્જુના સ્વાસ્થ્ય સમસ્યાઓ સુધી મર્યાદિત છે. વિષયની ચર્ચા કરવા માટે, કૃપા કરીને ડૉ. જીમેનેઝને પૂછો અથવા અમારો સંપર્ક કરો915-850-0900 .
ઘૂંટણની પીડા એ જાણીતું લક્ષણ છે જે ઘૂંટણની વિવિધ ઇજાઓ અને/અથવા પરિસ્થિતિઓને કારણે થઈ શકે છે, જેમાં�રમતો ઇજાઓ. ઘૂંટણ એ માનવ શરીરના સૌથી જટિલ સાંધાઓમાંનું એક છે કારણ કે તે ચાર હાડકાં, ચાર અસ્થિબંધન, વિવિધ રજ્જૂ, બે મેનિસ્કી અને કોમલાસ્થિના આંતરછેદથી બનેલું છે. અમેરિકન એકેડેમી ઓફ ફેમિલી ફિઝિશિયન્સ અનુસાર, ઘૂંટણની પીડાના સૌથી સામાન્ય કારણોમાં પેટેલર સબલક્સેશન, પેટેલર ટેન્ડિનિટિસ અથવા જમ્પર્સ ઘૂંટણ અને ઓસ્ગુડ-સ્લેટર રોગનો સમાવેશ થાય છે. જોકે ઘૂંટણનો દુખાવો 60 વર્ષથી વધુ ઉંમરના લોકોમાં થવાની સંભાવના છે, પરંતુ ઘૂંટણનો દુખાવો બાળકો અને કિશોરોમાં પણ થઈ શકે છે. ઘૂંટણના દુખાવાની સારવાર RICE પદ્ધતિઓને અનુસરીને ઘરે કરી શકાય છે, જો કે, ઘૂંટણની ગંભીર ઇજાઓને તાત્કાલિક તબીબી સહાયની જરૂર પડી શકે છે, જેમાં શિરોપ્રેક્ટિક સંભાળનો સમાવેશ થાય છે. �
ઓક્સિડેટીવ તણાવને મુક્ત રેડિકલ અથવા અસ્થિર પરમાણુઓ દ્વારા થતા કોષને થતા નુકસાન તરીકે વર્ણવવામાં આવે છે, જે આખરે તંદુરસ્ત કાર્યને અસર કરી શકે છે. માનવ શરીર બેક્ટેરિયા અને વાયરસને નિષ્ક્રિય કરવા માટે મુક્ત રેડિકલ બનાવે છે, જો કે, ઓક્સિજન, પ્રદૂષણ અને રેડિયેશન જેવા બાહ્ય પરિબળો ઘણીવાર મુક્ત રેડિકલ પણ ઉત્પન્ન કરી શકે છે. ઓક્સિડેટીવ તણાવ અસંખ્ય સ્વાસ્થ્ય સમસ્યાઓ સાથે સંકળાયેલ છે.
ઓક્સિડેટીવ તણાવ અને અન્ય તાણ આંતરિક રક્ષણાત્મક પદ્ધતિઓ ચાલુ કરે છે જે માનવ શરીરના એન્ટીઑકિસડન્ટ પ્રતિભાવને નિયંત્રિત કરવામાં મદદ કરી શકે છે. Nrf2 એ એક પ્રોટીન છે જે ઓક્સિડેટીવ તણાવના સ્તરને સમજે છે અને કોષોને આંતરિક અને બાહ્ય પરિબળોથી પોતાને બચાવવા માટે સક્ષમ બનાવે છે. એન્ટીઑકિસડન્ટ ઉત્સેચકો અને તાણ-પ્રતિભાવ જનીનોના ઉત્પાદનમાં સામેલ જનીનોને નિયંત્રિત કરવામાં મદદ કરવા માટે Nrf2 પણ દર્શાવવામાં આવ્યું છે. નીચેના લેખનો હેતુ સમજાવવાનો છે Nrf2 ની અસરો કેન્સર માં.
અમૂર્ત
Keap1-Nrf2 પાથવે ઓક્સિડેટીવ અને ઇલેક્ટ્રોફિલિક તણાવ માટે સાયટોપ્રોટેક્ટીવ પ્રતિભાવોનું મુખ્ય નિયમનકાર છે. જોકે ટ્રાન્સક્રિપ્શન ફેક્ટર Nrf2 દ્વારા ટ્રિગર થયેલા સેલ સિગ્નલિંગ પાથવે સામાન્ય અને પ્રિમેલિગ્નન્ટ પેશીઓમાં કેન્સરની શરૂઆત અને પ્રગતિને અટકાવે છે, સંપૂર્ણ રીતે જીવલેણ કોષોમાં Nrf2 પ્રવૃત્તિ કેન્સરના રસાયણ પ્રતિકારને વધારીને અને ગાંઠના કોષોની વૃદ્ધિને વધારીને વૃદ્ધિનો લાભ પૂરો પાડે છે. આ ગ્રાફિકલ સમીક્ષામાં, અમે Keap1-Nrf2 પાથવે અને કેન્સર કોષોમાં તેના ડિસરેગ્યુલેશનની ઝાંખી આપીએ છીએ. અમે કેન્સર કોશિકાઓમાં રચનાત્મક Nrf2 સક્રિયકરણના પરિણામો અને કેન્સર જીન થેરાપીમાં તેનો કેવી રીતે ઉપયોગ કરી શકાય છે તેનો પણ ટૂંકમાં સારાંશ આપીએ છીએ.
Keap1-Nrf2 પાથવે પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓ (ROS) અને ઇલેક્ટ્રોફાઇલ્સ [1] દ્વારા થતા અંતર્જાત અને બાહ્ય તણાવ માટે સાયટોપ્રોટેક્ટીવ પ્રતિભાવોનું મુખ્ય નિયમનકાર છે. પાથવેની અંદર મુખ્ય સિગ્નલિંગ પ્રોટીન ટ્રાન્સક્રિપ્શન ફેક્ટર Nrf2 (પરમાણુ પરિબળ એરિથ્રોઇડ 2-સંબંધિત પરિબળ 2) છે જે લક્ષ્ય જનીનોના નિયમનકારી પ્રદેશોમાં એન્ટીઑકિસડન્ટ પ્રતિભાવ તત્વ (ARE) સાથે નાના Maf પ્રોટીન સાથે જોડાય છે, અને Keap1 (Kelch ECH) એસોસિએટિંગ પ્રોટીન 1), એક દબાવનાર પ્રોટીન જે Nrf2 સાથે જોડાય છે અને યુબીક્વિટીન પ્રોટીઝોમ પાથવે (ફિગ. 1) દ્વારા તેના અધોગતિને પ્રોત્સાહન આપે છે. Keap1 એ ખૂબ જ સિસ્ટીન-સમૃદ્ધ પ્રોટીન છે, માઉસ Keap1 કુલ 25 અને માનવીય 27 સિસ્ટીન અવશેષો ધરાવે છે, જેમાંથી મોટા ભાગનાને વિટ્રોમાં વિવિધ ઓક્સિડન્ટ્સ અને ઇલેક્ટ્રોફાઈલ્સ [2] દ્વારા સુધારી શકાય છે. આમાંના ત્રણ અવશેષો, C151, C273 અને C288, Keap1 ની રચનામાં ફેરફાર કરીને Nrf2 અને અનુગામી લક્ષ્ય જનીન અભિવ્યક્તિ [3] (ફિગ. 1) તરફ દોરી જતા કાર્યાત્મક ભૂમિકા ભજવતા દર્શાવવામાં આવ્યા છે. ચોક્કસ પદ્ધતિ કે જેના દ્વારા Keap1 માં સિસ્ટીન ફેરફારો Nrf2 સક્રિયકરણ તરફ દોરી જાય છે તે જાણી શકાયું નથી, પરંતુ બે પ્રચલિત પરંતુ પરસ્પર વિશિષ્ટ મોડલ છે (1) હિંગ અને લેચ મોડલ, જેમાં Keap1 ના IVR માં રહેતા થિયોલ અવશેષોમાં Keap1 ફેરફારો થાય છે. Nrf2 સાથેની ક્રિયાપ્રતિક્રિયાને વિક્ષેપિત કરે છે જે Nrf2 ની અંદરના લાયસિન અવશેષોની ખોટી ગોઠવણીનું કારણ બને છે જે લાંબા સમય સુધી પોલીયુબીક્વિટીનિલેટેડ હોઈ શકતું નથી અને (2) મોડેલ કે જેમાં થિયોલ ફેરફાર Cul3 ને Keap1 [3] થી અલગ કરવાનું કારણ બને છે. બંને મોડલમાં, પ્રેરક-સંશોધિત અને Nrf2-બાઉન્ડ Keap1 નિષ્ક્રિય છે અને પરિણામે, નવા સંશ્લેષિત Nrf2 પ્રોટીન કેપ1ને બાયપાસ કરે છે અને ન્યુક્લિયસમાં સ્થાનાંતરિત થાય છે, ARE સાથે જોડાય છે અને NAD(P)H જેવા Nrf2 લક્ષ્ય જનીનોની અભિવ્યક્તિ ચલાવે છે. ક્વિનોન oxidoreductase 1 (NQO1), heme oxygenase 1 (HMOX1), glutamate-cysteine ligase (GCL) અને glutathione S transferases (GSTs) (ફિગ. 2). Nrf1 લક્ષ્ય જનીન ઇન્ડક્શનમાં પરિણમતા Keap2 થિઓલ્સના ફેરફારો ઉપરાંત, p21 અને p62 જેવા પ્રોટીન Nrf2 અથવા Keap1 સાથે જોડાઈ શકે છે જેથી Nrf2 અને Keap1 [1], [3] (ફિગ. 3) વચ્ચેની ક્રિયાપ્રતિક્રિયામાં વિક્ષેપ પડે છે.
ફિગ. 1. Nrf2 અને Keap1 અને સિસ્ટીન કોડનું માળખું. (A) Nrf2 માં 589 એમિનો એસિડનો સમાવેશ થાય છે અને તેમાં છ ઉત્ક્રાંતિપૂર્વક અત્યંત સંરક્ષિત ડોમેન્સ છે, Neh1-6. Neh1 એક bZip મોટિફ ધરાવે છે, એક મૂળભૂત ક્ષેત્ર � લ્યુસીન ઝિપર (L-Zip) માળખું, જ્યાં મૂળભૂત પ્રદેશ DNA ઓળખ માટે જવાબદાર છે અને L-Zip નાના Maf પ્રોટીન સાથે ડાઇમરાઇઝેશનની મધ્યસ્થી કરે છે. Neh6 ન્યુક્લિયસમાં Nrf2 ના અધોગતિને મધ્યસ્થી કરવા માટે ડિગ્રોન તરીકે કાર્ય કરે છે. Neh4 અને 5 ટ્રાન્સએક્ટિવેશન ડોમેન્સ છે. Neh2 માં ETGE અને DLG મોટિફ્સ છે, જે Keap1 સાથે ક્રિયાપ્રતિક્રિયા માટે જરૂરી છે, અને lysine અવશેષો (7 K), જે Keap1-આશ્રિત પોલીયુબીક્વિટીનેશન અને Nrf2 ના અધોગતિ માટે અનિવાર્ય છે. (B) Keap1 માં 624 એમિનો એસિડ અવશેષો છે અને તેમાં પાંચ ડોમેન્સ છે. બે પ્રોટીન-પ્રોટીન ક્રિયાપ્રતિક્રિયાના હેતુઓ, BTB ડોમેન અને કેલ્ચ ડોમેન, મધ્યસ્થી પ્રદેશ (IVR) દ્વારા અલગ પડે છે. IVR ના N-ટર્મિનલ ભાગ સાથે BTB ડોમેન, Keap1 ના હોમોડિમેરાઇઝેશન અને Cullin3 (Cul3) સાથે બંધનમાં મધ્યસ્થી કરે છે. કેલ્ચ ડોમેન અને સી-ટર્મિનલ પ્રદેશ Neh2 સાથેની ક્રિયાપ્રતિક્રિયામાં મધ્યસ્થી કરે છે. (C) Nrf2 તેના Neh1 ETGE અને DLG મોટિફ્સ દ્વારા Keap2 ના બે અણુઓ સાથે ક્રિયાપ્રતિક્રિયા કરે છે. ETGE અને DLG બંને Keap1 કેલ્ચ મોટિફની નીચેની સપાટી પર સમાન સાઇટ્સ સાથે જોડાય છે. (D) Keap1 માનવ પ્રોટીનમાં 27 સિસ્ટીન સાથે સિસ્ટીન અવશેષોથી સમૃદ્ધ છે. આમાંના કેટલાક સિસ્ટીન મૂળભૂત અવશેષોની નજીક સ્થિત છે અને તેથી ઇલેક્ટ્રોફાઇલ્સ અને ઓક્સિડન્ટ્સનું ઉત્તમ લક્ષ્ય છે. ઇલેક્ટ્રોફાઇલ્સ દ્વારા સિસ્ટીન અવશેષોના ફેરફારની પેટર્નને સિસ્ટીન કોડ તરીકે ઓળખવામાં આવે છે. સિસ્ટીન કોડની પૂર્વધારણા સૂચવે છે કે માળખાકીય રીતે અલગ Nrf2 એક્ટિવેટીંગ એજન્ટો વિવિધ Keap1 સિસ્ટીનને અસર કરે છે. સિસ્ટીન ફેરફારો Nrf1 DLG અને Keap2 કેલ્ચ ડોમેન્સ વચ્ચેની ક્રિયાપ્રતિક્રિયામાં વિક્ષેપ પાડતા Keap1 માં રચનાત્મક ફેરફારો તરફ દોરી જાય છે, આમ Nrf2 ની બહુયુબિક્વિટીનેશનને અટકાવે છે. Cys151, Cys273 અને Cys288 નું કાર્યાત્મક મહત્વ દર્શાવવામાં આવ્યું છે, કારણ કે ઇન્ડ્યુસર્સ [273], [288] દ્વારા Nrf2 ને સક્રિય કરવા માટે Nrf151 અને Cys2 ને દબાવવા માટે Cys1 અને Cys3 જરૂરી છે.
ફિગ. 2. Nrf2-Keap1 સિગ્નલિંગ પાથવે. (A અને B) મૂળભૂત પરિસ્થિતિઓમાં, બે Keap1 પરમાણુ Nrf2 અને Nrf2 સાથે જોડાય છે Cul3-આધારિત E3 ligase કોમ્પ્લેક્સ દ્વારા બહુવ્યાપક છે. આ પોલીયુબીક્વિટીલેશન પ્રોટીઝોમ દ્વારા ઝડપી Nrf2 અધોગતિમાં પરિણમે છે. Nrf2 નો એક નાનો હિસ્સો અવરોધક સંકુલમાંથી છટકી જાય છે અને મૂળભૂત ARE-આશ્રિત જનીન અભિવ્યક્તિને મધ્યસ્થી કરવા માટે ન્યુક્લિયસમાં એકઠા થાય છે, ત્યાં સેલ્યુલર હોમિયોસ્ટેસિસ જાળવી રાખે છે. (C) તાણની પરિસ્થિતિઓમાં, પ્રેરકો કેપ1 સિસ્ટીનમાં ફેરફાર કરે છે જે અવરોધક સંકુલના વિયોજન દ્વારા Nrf2 સર્વવ્યાપકતાના અવરોધ તરફ દોરી જાય છે. (D) મિજાગરું અને લેચ મોડલ મુજબ, ચોક્કસ Keap1 સિસ્ટીન અવશેષોમાં ફેરફાર કરવાથી Keap1 માં રચનાત્મક ફેરફારો થાય છે જેના પરિણામે Keap2 થી Nrf1 DLG મોટિફને અલગ કરવામાં આવે છે. Nrf2 ની સર્વવ્યાપકતા વિક્ષેપિત છે પરંતુ ETGE મોટિફ સાથે બંધન રહે છે. (E) Keap1-Cul3 ડિસોસિએશન મોડેલમાં, Keap1 અને Cul3 નું બંધન ઇલેક્ટ્રોફાઇલ્સના પ્રતિભાવમાં વિક્ષેપિત થાય છે, જે સર્વવ્યાપક પદ્ધતિમાંથી Nrf2 ના ભાગી જવા તરફ દોરી જાય છે. સૂચવેલા બંને મોડેલોમાં, પ્રેરક-સંશોધિત અને Nrf2-બાઉન્ડ Keap1 નિષ્ક્રિય છે અને પરિણામે, નવા સંશ્લેષિત Nrf2 પ્રોટીન કેપ1ને બાયપાસ કરે છે અને ન્યુક્લિયસમાં સ્થાનાંતરિત થાય છે, એન્ટિઓક્સિડન્ટ રિસ્પોન્સ એલિમેન્ટ (ARE) સાથે જોડાય છે અને Nrf2 લક્ષ્યની અભિવ્યક્તિને ચલાવે છે. જનીનો જેમ કે NQO1, HMOX1, GCL અને GST [1], [3].
ફિગ. 3. કેન્સરમાં Nrf2 ના રચનાત્મક પરમાણુ સંચય માટેની પદ્ધતિઓ. (A) Nrf2 અથવા Keap1 માં સોમેટિક મ્યુટેશન આ બે પ્રોટીનની ક્રિયાપ્રતિક્રિયાને વિક્ષેપિત કરે છે. Nrf2 માં, પરિવર્તનો ETGE અને DLG મોટિફ્સને અસર કરે છે, પરંતુ Keap1 માં પરિવર્તન વધુ સમાનરૂપે વિતરિત થાય છે. વધુમાં, ઓન્કોજીન સક્રિયકરણ, જેમ કે KrasG12D[5], અથવા PTEN [11] જેવા ટ્યુમર સપ્રેસર્સમાં વિક્ષેપ, Nrf2 ના ટ્રાન્સક્રિપ્શનલ ઇન્ડક્શન અને ન્યુક્લિયર Nrf2 માં વધારો તરફ દોરી શકે છે. (B) ફેફસાં અને પ્રોસ્ટેટ કેન્સરમાં Keap1 પ્રમોટરનું હાઇપરમેથિલેશન Keap1 mRNA અભિવ્યક્તિમાં ઘટાડો તરફ દોરી જાય છે, જે Nrf2 [6], [7] ના પરમાણુ સંચયમાં વધારો કરે છે. (C) પારિવારિક પેપિલરી રેનલ કાર્સિનોમામાં, ફ્યુમરેટ હાઇડ્રેટેજ એન્ઝાઇમ પ્રવૃત્તિનું નુકસાન ફ્યુમરેટના સંચય તરફ દોરી જાય છે અને આગળ Keap1 સિસ્ટીન અવશેષો (2SC) ના સક્સીનેશન તરફ દોરી જાય છે. આ પોસ્ટ-ટ્રાન્સલેશનલ ફેરફાર Keap1-Nrf2 ક્રિયાપ્રતિક્રિયામાં વિક્ષેપ અને Nrf2 [8], [9] ના પરમાણુ સંચય તરફ દોરી જાય છે. (D) p62 અને p21 જેવા વિક્ષેપકર્તા પ્રોટીનનું સંચય Nrf2-Keap1 બંધનને ખલેલ પહોંચાડી શકે છે અને પરિણામે પરમાણુ Nrf2 માં વધારો થાય છે. p62 Nrf1 માટે બંધાઈ રહેલા પોકેટને ઓવરલેપ કરીને Keap2 સાથે જોડાય છે અને p21 Nrf2 ના DLG અને ETGE મોટિફ્સ સાથે સીધો સંપર્ક કરે છે, ત્યાંથી Keap1 [10] સાથે સ્પર્ધા કરે છે.
કેન્સરમાં Nrf2 ના સક્રિયકરણ અને ડિસરેગ્યુલેશનની પદ્ધતિઓ
Nrf2 સક્રિયકરણ દ્વારા પ્રદાન કરવામાં આવેલ સાયટોપ્રોટેક્શન સામાન્ય અને પ્રિમેલિગ્નન્ટ પેશીઓમાં કેન્સર કેમોપ્રિવેન્શન માટે મહત્વપૂર્ણ છે, તેમ છતાં, સંપૂર્ણપણે જીવલેણ કોષોમાં Nrf2 પ્રવૃત્તિ કેન્સરના રસાયણ પ્રતિકારને વધારીને અને ટ્યુમર સેલ વૃદ્ધિ [4] વધારીને વૃદ્ધિનો લાભ પ્રદાન કરે છે. કેટલીક પદ્ધતિઓ કે જેના દ્વારા Nrf2 સિગ્નલિંગ પાથવે વિવિધ કેન્સરમાં રચનાત્મક રીતે સક્રિય થાય છે તેનું વર્ણન કરવામાં આવ્યું છે: (1) Keap1 માં સોમેટિક મ્યુટેશન અથવા Nrf1 ના Keap2 બંધનકર્તા ડોમેન તેમની ક્રિયાપ્રતિક્રિયામાં ખલેલ પહોંચાડે છે; (2) Keap1 અભિવ્યક્તિનું એપિજેનેટિક સાયલન્સિંગ Nrf2 ના ખામીયુક્ત દમન તરફ દોરી જાય છે; (3) વિક્ષેપકર્તા પ્રોટીનનું સંચય જેમ કે p62 કેપ1-Nrf2 સંકુલના વિયોજન તરફ દોરી જાય છે; (4) ઓન્કોજેનિક K-Ras, B-Raf અને c-Myc દ્વારા Nrf2 નું ટ્રાન્સક્રિપ્શનલ ઇન્ડક્શન; અને (5) ફ્યુમરેટ હાઇડ્રેટેજ એન્ઝાઇમ પ્રવૃત્તિ [1], [3], [4], [5], [6], [7], [8], [9], [10], [3], [2], [2] 4], [2], [11] (ફિગ. 4). બંધારણીય રીતે વિપુલ પ્રમાણમાં NrfXNUMX પ્રોટીન દવાના ચયાપચયમાં સામેલ જનીનોની અભિવ્યક્તિમાં વધારો કરે છે જેનાથી કીમોથેરાપ્યુટિક દવાઓ અને રેડિયોથેરાપી સામે પ્રતિકાર વધે છે. વધુમાં, ઉચ્ચ NrfXNUMX પ્રોટીન સ્તર કેન્સર [XNUMX] માં નબળા પૂર્વસૂચન સાથે સંકળાયેલું છે. ઓવરએક્ટિવ NrfXNUMX એ ગ્લુકોઝ અને ગ્લુટામાઇનને એનાબોલિક માર્ગો તરફ નિર્દેશિત કરીને પ્યુરિન સંશ્લેષણમાં વધારો કરીને અને કોષના પ્રસારને પ્રોત્સાહન આપવા પેન્ટોઝ ફોસ્ફેટ માર્ગને પ્રભાવિત કરીને પણ અસર કરે છે [XNUMX] (ફિગ. XNUMX).
ફિગ. 4. ટ્યુમોરીજેનેસિસમાં Nrf2 ની બેવડી ભૂમિકા. શારીરિક પરિસ્થિતિઓ હેઠળ, ન્યુક્લિયર Nrf2 નું નીચું સ્તર સેલ્યુલર હોમિયોસ્ટેસિસની જાળવણી માટે પૂરતું છે. Nrf2 કાર્સિનોજેન્સ, ROS અને અન્ય DNA-નુકસાનકર્તા એજન્ટોને દૂર કરીને ગાંઠની શરૂઆત અને કેન્સર મેટાસ્ટેસિસને અટકાવે છે. ટ્યુમોરીજેનેસિસ દરમિયાન, ડીએનએ નુકસાન એકઠા થવાથી Nrf2 ની રચનાત્મક હાયપરએક્ટિવિટી થાય છે જે સ્વાયત્ત જીવલેણ કોષોને અંતર્જાત આરઓએસના ઉચ્ચ સ્તરને સહન કરવામાં અને એપોપ્ટોસિસને ટાળવામાં મદદ કરે છે. સતત એલિવેટેડ ન્યુક્લિયર Nrf2 સ્તર મેટાબોલિક રિપ્રોગ્રામિંગ અને ઉન્નત સેલ પ્રસારમાં ફાળો આપતા સાયટોપ્રોટેક્ટીવ જનીનો ઉપરાંત મેટાબોલિક જનીનોને સક્રિય કરે છે. ઉચ્ચ Nrf2 સ્તર ધરાવતા કેન્સર રેડિયો અને કેમોરેસિસ્ટન્સ અને આક્રમક કેન્સર સેલ પ્રસારને કારણે નબળા પૂર્વસૂચન સાથે સંકળાયેલા છે. આમ, Nrf2 પાથવે પ્રવૃત્તિ ટ્યુમોરીજેનેસિસના પ્રારંભિક તબક્કામાં રક્ષણાત્મક છે, પરંતુ પછીના તબક્કામાં નુકસાનકારક છે. તેથી, કેન્સરની રોકથામ માટે, Nrf2 પ્રવૃત્તિને વધારવી એ એક મહત્વપૂર્ણ અભિગમ છે જ્યારે કેન્સરની સારવાર માટે, Nrf2 નિષેધ ઇચ્છનીય છે [4], [11].
પ્રતિકૂળ પરિણામો સાથે કેન્સર કોષોમાં સામાન્ય રીતે ઉચ્ચ Nrf2 પ્રવૃત્તિ જોવા મળે છે તે જોતાં, Nrf2 ને રોકવા માટે ઉપચારની જરૂર છે. કમનસીબે, કેટલાક અન્ય bZip પરિવારના સભ્યો સાથે માળખાકીય સમાનતાને લીધે, ચોક્કસ Nrf2 અવરોધકોનો વિકાસ એ એક પડકારજનક કાર્ય છે અને Nrf2 નિષેધના માત્ર થોડા અભ્યાસો જ આજ સુધી પ્રકાશિત કરવામાં આવ્યા છે. કુદરતી ઉત્પાદનોની તપાસ કરીને, રેન એટ અલ. [૧૨] એન્ટિનોપ્લાસ્ટિક સંયોજન બ્રુસાટોલને Nrf12 અવરોધક તરીકે ઓળખવામાં આવે છે જે સિસ્પ્લેટિનની કીમોથેરાપ્યુટિક અસરકારકતાને વધારે છે. વધુમાં, PI2K અવરોધકો [3], [11] અને Nrf13 siRNA [2] નો ઉપયોગ કેન્સરના કોષોમાં Nrf14 ને રોકવા માટે કરવામાં આવ્યો છે. તાજેતરમાં, અમે એક વૈકલ્પિક અભિગમનો ઉપયોગ કર્યો છે, જેને કેન્સર સુસાઈડ જીન થેરાપી તરીકે ઓળખવામાં આવે છે, ઉચ્ચ Nrf2 સ્તરો ધરાવતા કેન્સર કોષોને લક્ષ્ય બનાવવા માટે. Nrf2-સંચાલિત લેન્ટીવાયરલ વેક્ટર [2] જેમાં thymidine kinase (TK) હોય છે તે ઉચ્ચ ARE પ્રવૃત્તિ સાથે કેન્સરના કોષોમાં સ્થાનાંતરિત થાય છે અને કોષોને પ્રો-ડ્રગ, ગેન્સીક્લોવીર (GCV) વડે સારવાર આપવામાં આવે છે. GCV ને GCV-મોનોફોસ્ફેટમાં ચયાપચય કરવામાં આવે છે, જે સેલ્યુલર કિનાસીસ દ્વારા ઝેરી ટ્રાઇફોસ્ફેટ સ્વરૂપમાં ફોસ્ફોરીલેટેડ થાય છે [15] (ફિગ. 16). આનાથી માત્ર ટ્યુમર કોશિકાઓ ધરાવતા TK જ નહીં, પરંતુ બાયસ્ટેન્ડર અસર [5]ને કારણે પડોશી કોષોને પણ અસરકારક રીતે મારવામાં આવે છે. ARE-રેગ્યુલેટેડ TK/GCV જીન થેરાપીને કેન્સરના કીમોથેરાપ્યુટિક એજન્ટ ડોક્સોરુબીસીનને સારવારમાં જોડીને વધુ વધારી શકાય છે [17], આ વિચારને સમર્થન આપે છે કે આ અભિગમ પરંપરાગત ઉપચારો સાથે જોડાણમાં ઉપયોગી થઈ શકે છે.
ફિગ. 5. આત્મઘાતી જનીન ઉપચાર. કેન્સરના કોષોમાં બંધારણીય Nrf2 પરમાણુ સંચયનું કેન્સર આત્મહત્યા જનીન ઉપચાર [2] માટે Nrf16-ચાલિત વાયરલ વેક્ટરનો ઉપયોગ કરીને શોષણ કરી શકાય છે. આ અભિગમમાં, ચાર ARE સાથે ન્યૂનતમ SV40 પ્રમોટર હેઠળ થાઇમિડિન કિનેઝ (TK) ને વ્યક્ત કરતું લેન્ટીવાયરલ વેક્ટર (LV) ફેફસાના એડેનોકાર્સિનોમા કોષોમાં પરિવર્તિત થાય છે. ઉચ્ચ પરમાણુ Nrf2 સ્તરો Nrf2 બંધનકર્તા દ્વારા TK ની મજબૂત અભિવ્યક્તિ તરફ દોરી જાય છે. ત્યારબાદ કોષોને પ્રો-ડ્રગ, ગેન્સીક્લોવીર (જીસીવી) વડે સારવાર આપવામાં આવે છે, જે ટીકે દ્વારા ફોસ્ફોરીલેટેડ છે. ટ્રાઇફોસ્ફોરીલેટેડ જીસીવી ડીએનએ સંશ્લેષણને વિક્ષેપિત કરે છે અને બાયસ્ટેન્ડર અસરને કારણે માત્ર ટીકે ધરાવતા ટ્યુમર કોષોને જ નહીં, પરંતુ પડોશી કોષોને પણ અસરકારક રીતે મારવા તરફ દોરી જાય છે.
Nrf2 એક માસ્ટર રેગ્યુલેટર છે જે માનવ શરીરમાં શક્તિશાળી એન્ટીઑકિસડન્ટોના ઉત્પાદનને ટ્રિગર કરે છે જે ઓક્સિડેટીવ તણાવને દૂર કરવામાં મદદ કરે છે. વિવિધ એન્ટીઑકિસડન્ટ ઉત્સેચકો, જેમ કે સુપરઓક્સાઇડ ડિસમ્યુટેઝ, અથવા SOD, ગ્લુટાથિઓન અને કેટાલેઝ, પણ Nrf2 માર્ગ દ્વારા સક્રિય થાય છે. વધુમાં, હળદર, અશ્વગંધા, બેકોપા, ગ્રીન ટી અને મિલ્ક થીસલ જેવા અમુક ફાયટોકેમિકલ્સ Nrf2 ને સક્રિય કરે છે. સંશોધન અભ્યાસમાં જાણવા મળ્યું છે કે Nrf2 સક્રિયકરણ કુદરતી રીતે સેલ્યુલર સંરક્ષણને વધારી શકે છે અને માનવ શરીરમાં સંતુલન પુનઃસ્થાપિત કરી શકે છે.
ડૉ. એલેક્સ જિમેનેઝ ડીસી, CCST ઇનસાઇટ
સલ્ફોરાફેન અને કેન્સર, મૃત્યુદર, વૃદ્ધત્વ, મગજ અને વર્તન, હૃદય રોગ અને વધુ પર તેની અસરો
Isothiocyanates એ કેટલાક સૌથી મહત્વપૂર્ણ છોડ સંયોજનો છે જે તમે તમારા આહારમાં મેળવી શકો છો. આ વિડિયોમાં હું તેમના માટે અત્યાર સુધીનો સૌથી વ્યાપક કેસ બનાવું છું. ટૂંકા ધ્યાન ગાળો? નીચેના ટાઈમ પોઈન્ટ્સમાંથી કોઈ એક પર ક્લિક કરીને તમારા મનપસંદ વિષય પર જાઓ. નીચે સંપૂર્ણ સમયરેખા.
મુખ્ય વિભાગો:
00:01:14 - કેન્સર અને મૃત્યુદર
00:19:04 - વૃદ્ધત્વ
00:26:30 - મગજ અને વર્તન
00:38:06 - અંતિમ રીકેપ
00:40:27 - માત્રા
સંપૂર્ણ સમયરેખા:
00:00:34 - સલ્ફોરાફેનનો પરિચય, વિડીયોનું મુખ્ય ધ્યાન.
00:01:14 - ક્રુસિફેરસ શાકભાજીનો વપરાશ અને સર્વ-કારણ મૃત્યુદરમાં ઘટાડો.
00:44:24 - શાકભાજીમાંથી સક્રિય માયરોસિનેઝ સાથે જોડવામાં આવે ત્યારે પૂરક વધુ સારી રીતે કામ કરે છે.
00:44:56 - રાંધવાની તકનીકો અને ક્રુસિફેરસ શાકભાજી.
00:46:06 - ગોઇટ્રોજન તરીકે આઇસોથિયોસાયનેટ્સ.
સમર્થન
આ કાર્યને ફિનલેન્ડની એકેડેમી, સિગ્રિડ જ્યુસેલિયસ ફાઉન્ડેશન અને ફિનિશ કેન્સર સંસ્થાઓ દ્વારા સમર્થન આપવામાં આવ્યું હતું.
નિષ્કર્ષમાં, ન્યુક્લિયર ફેક્ટર (એરિથ્રોઇડ-ડેરિવ્ડ 2)-જેવું 2, જેને NFE2L2 અથવા Nrf2 તરીકે પણ ઓળખવામાં આવે છે, તે એક પ્રોટીન છે જે એન્ટીઑકિસડન્ટોના ઉત્પાદનમાં વધારો કરે છે જે માનવ શરીરને ઓક્સિડેટીવ તણાવ સામે રક્ષણ આપે છે. ઉપર વર્ણવ્યા મુજબ, Nrf2 પાથવેની ઉત્તેજના કેન્સર સહિત ઓક્સિડેટીવ તણાવને કારણે થતા રોગોની સારવાર માટે અભ્યાસ કરવામાં આવી રહી છે. અમારી માહિતીનો અવકાશ શિરોપ્રેક્ટિક અને કરોડરજ્જુના સ્વાસ્થ્ય સમસ્યાઓ સુધી મર્યાદિત છે. વિષયની ચર્ચા કરવા માટે, કૃપા કરીને ડૉ. જીમેનેઝને પૂછો અથવા અમારો સંપર્ક કરો915-850-0900 .
ઘૂંટણની પીડા એ જાણીતું લક્ષણ છે જે ઘૂંટણની વિવિધ ઇજાઓ અને/અથવા પરિસ્થિતિઓને કારણે થઈ શકે છે, જેમાં�રમતો ઇજાઓ. ઘૂંટણ એ માનવ શરીરના સૌથી જટિલ સાંધાઓમાંનું એક છે કારણ કે તે ચાર હાડકાં, ચાર અસ્થિબંધન, વિવિધ રજ્જૂ, બે મેનિસ્કી અને કોમલાસ્થિના આંતરછેદથી બનેલું છે. અમેરિકન એકેડેમી ઓફ ફેમિલી ફિઝિશિયન્સ અનુસાર, ઘૂંટણની પીડાના સૌથી સામાન્ય કારણોમાં પેટેલર સબલક્સેશન, પેટેલર ટેન્ડિનિટિસ અથવા જમ્પર્સ ઘૂંટણ અને ઓસ્ગુડ-સ્લેટર રોગનો સમાવેશ થાય છે. જોકે ઘૂંટણનો દુખાવો 60 વર્ષથી વધુ ઉંમરના લોકોમાં થવાની સંભાવના છે, પરંતુ ઘૂંટણનો દુખાવો બાળકો અને કિશોરોમાં પણ થઈ શકે છે. ઘૂંટણના દુખાવાની સારવાર RICE પદ્ધતિઓને અનુસરીને ઘરે કરી શકાય છે, જો કે, ઘૂંટણની ગંભીર ઇજાઓને તાત્કાલિક તબીબી સારવારની જરૂર પડી શકે છે, જેમાં શિરોપ્રેક્ટિક સંભાળનો સમાવેશ થાય છે.
ડીએનએ આશરે 20,000 જનીનોને સમર્થન આપે છે, દરેકમાં તંદુરસ્ત જીવનશૈલી માટે જરૂરી પ્રોટીન અથવા એન્ઝાઇમ બનાવવા માટેનો પ્રોગ્રામ હોય છે. આમાંની દરેક પેટર્નને એક પ્રકારનાં "પ્રમોટર" દ્વારા સતત નિયંત્રિત કરવાની જરૂર છે જે દરેક પદાર્થ અને/અથવા રસાયણમાંથી કેટલું જનરેટ થાય છે અને તે કઈ પરિસ્થિતિઓમાં વિકસિત થશે તેનું બરાબર સંચાલન કરે છે.
એન્ટીઑકિસડન્ટ રિસ્પોન્સ એલિમેન્ટ અથવા ARE તરીકે ઓળખાતા સ્વિચ જેવા પ્રમોટર વિસ્તારોના ચોક્કસ પ્રકાર સાથે કનેક્ટ કરીને, Nrf2 પરિબળસેંકડો અલગ જનીનો માટે સર્જનની ઝડપને સમર્થન આપે છે જે કોષોને તણાવપૂર્ણ સંજોગોમાં ટકી રહેવા સક્ષમ બનાવે છે. આ જનીનો પછી એન્ટીઑકિસડન્ટ ઉત્સેચકોની પસંદગી ઉત્પન્ન કરે છે જે ઓક્સિડન્ટને તટસ્થ કરીને અને તેમના ઉત્પાદનમાં પાછળ રહી ગયેલા ઝેરી ઉપ-ઉત્પાદનોને સાફ કરીને સંરક્ષણ નેટવર્ક વિકસાવે છે, ઉપરાંત તેઓને થયેલા નુકસાનને પુનઃસ્થાપિત કરવામાં મદદ કરે છે.
ઓક્સિડેટીવ સ્ટ્રેસ શું છે?
સુપરઓક્સાઇડ રેડિકલ, અથવા O2-., અને હાઇડ્રોજન પેરોક્સાઇડ, અથવા H2O2 જેવા કેટલાક ઓક્સિડન્ટ્સ માનવ શરીરને ટકાવી રાખતા પદાર્થો અને/અથવા રસાયણોને બાળી નાખવાની પ્રથા દ્વારા બનાવવામાં આવ્યા છે. માનવ શરીરમાં એન્ટીઑકિસડન્ટ ઉત્સેચકો હોય છે જે અમે જે પ્રતિક્રિયાશીલ ખોરાક અને પીણાંનો વપરાશ કરીએ છીએ તેને તટસ્થ અને ડિટોક્સિફાય કરે છે. Nrf2 સંતુલન જાળવવા માટે તેમના ઉત્પાદનને મોડ્યુલેટ કરે છે અને આ તમામ ઉત્સેચકોની માંગને અન્ડરસ્કોર કરે છે. આ સંતુલન વય સહિતના કેટલાક પરિબળો દ્વારા વિક્ષેપિત થઈ શકે છે.
જેમ જેમ આપણે વય કરીએ છીએ તેમ, માનવ શરીર ઓછું Nrf2 બનાવે છે અને આ નાજુક સંતુલન ધીમે ધીમે ઓક્સિડેટીવ બાજુ તરફ વળવાનું શરૂ કરી શકે છે, જે સ્થિતિને ઓક્સિડેટીવ તણાવ તરીકે ઓળખવામાં આવે છે. રોગ ઓક્સિડન્ટ્સનું વધુ પડતું ઉત્પાદન પણ કરી શકે છે. ચેપ, એલર્જી અને સ્વયંપ્રતિરક્ષા વિકૃતિઓ ઉપરાંત O2- જેવા પ્રતિક્રિયાશીલ ઓક્સિડન્ટ્સ બનાવવા માટે આપણા રોગપ્રતિકારક કોષોને ટ્રિગર કરી શકે છે. , H2O2, OH અને HOCl, જ્યાં તંદુરસ્ત કોષો ક્ષતિગ્રસ્ત થાય છે અને બળતરા સાથે પ્રતિક્રિયા આપે છે. વૃદ્ધત્વ સાથે સંકળાયેલ રોગો, જેમાં હૃદયરોગનો હુમલો, સ્ટ્રોક, કેન્સર, અને અલ્ઝાઈમર રોગ જેવી ન્યુરોડીજનરેટિવ સ્થિતિઓ પણ ઓક્સિડન્ટ્સના વિકાસમાં વધારો કરે છે, તણાવ પેદા કરે છે અને બળતરા પ્રતિભાવમાં વધારો કરે છે.
Nrf2 એક્ટિવેટર્સ શું છે?
Nrf2 પ્રોટીન, જેને તે ઉત્સેચકો અને જનીનોને ટેકો અને નિયંત્રિત કરી શકે છે તેના કારણે ટ્રાન્સક્રિપ્શન પરિબળ પણ કહેવાય છે, તે કોષની અંદર બાયોકેમિકલ પ્રતિક્રિયાઓના ક્રમનું ગુપ્ત તત્વ છે જે જ્ઞાનાત્મક સંતુલન તેમજ ઓક્સિડેટીવ સંતુલનમાં ફેરફાર પર પ્રતિક્રિયા આપે છે. આ પાથવેના સંવેદના તત્વો Nrf2 ને સંશોધિત કરે છે અને વિસર્જિત કરે છે, તેને ટ્રિગર કરે છે જેથી તે કોષના ન્યુક્લિયસમાં DNA તરફ ફેલાઈ શકે. Nrf2 વૈકલ્પિક રીતે જનીનો અને ઉત્સેચકોને ચાલુ અથવા બંધ કરી શકે છે જેને તે કોષને સુરક્ષિત કરવા માટે સપોર્ટ કરે છે.
સદનસીબે, Nrf2 એક્ટિવેટર્સ એવા વિવિધ પદાર્થોનો વિકાસ સદીઓ પહેલા ચાઈનીઝ અને નેટિવ અમેરિકન પરંપરાગત ઉપાયોમાં અમુક છોડ અને અર્કના વપરાશ દ્વારા થાય છે. આ ફાયટોકેમિકલ્સ ઓછા આડ-અસર સાથે એટલા જ શક્તિશાળી લાગે છે, જેમ કે Nrf2-સક્રિય કરતી ફાર્માસ્યુટિકલ પ્રોડક્ટ્સ જે આજે ઉપયોગમાં લેવાય છે.
ન્યુક્લિયર ફેક્ટર એરિથ્રોઇડ 2-સંબંધિત પરિબળ, જે સામાન્ય રીતે Nrf2 તરીકે ઓળખાય છે, તે એક ટ્રાન્સક્રિપ્શન પરિબળ છે જે જનીનો, ઉત્સેચકો અને એન્ટીઑકિસડન્ટ પ્રતિક્રિયાઓનું નિયમન કરીને કોષનું રક્ષણ કરે છે. ટ્રાન્સક્રિપ્શન પરિબળો એ પ્રોટીનનો એક પ્રકાર છે જે ગ્લુટાથિઓન એસ-ટ્રાન્સફેરેસ અથવા GST સહિત ચોક્કસ પદાર્થો અને રસાયણોના નિર્માણને પ્રોત્સાહન આપવા માટે ડીએનએ સાથે જોડાય છે. Nrf2 સક્રિયકરણ સક્રિય પ્રોટીનના ઉત્પાદનને પ્રેરિત કરે છે જે ઓક્સિડેટીવ તણાવ ઘટાડવામાં મદદ કરવા માટે શક્તિશાળી એન્ટીઑકિસડન્ટ ક્ષમતા દર્શાવે છે.
ડૉ. એલેક્સ જિમેનેઝ ડીસી, CCST ઇનસાઇટ
Nrf2 સક્રિયકરણ પાછળનું વિજ્ઞાન
એકવાર પ્રારંભિક Nrf2-સક્રિય આહાર પૂરક 2004 માં બનાવવામાં આવ્યા પછી, Nrf2 પાથવેના કાર્ય વિશે ન્યૂનતમ માહિતી જાણીતી હતી. Nrf200 પરના સાહિત્યમાં આશરે 2 અખબારો, જેને પરમાણુ પરિબળ-જેવા 2 અથવા NFE2L2 તરીકે પણ ઓળખવામાં આવે છે, અસ્તિત્વમાં છે અને સંશોધકો માત્ર સસ્તન પ્રાણીઓમાં Nrf2 ના એન્ટીઑકિસડન્ટ પ્રતિભાવ શોધવાનું શરૂ કરી રહ્યા હતા. 2017 સુધીમાં, જો કે, આ “માસ્ટર રેગ્યુલેટર” પર 9,300 થી વધુ શૈક્ષણિક સંશોધન અભ્યાસો છાપવામાં આવ્યા છે.
વાસ્તવમાં, Nrf2 ઘણા એન્ટીઑકિસડન્ટ ઉત્સેચકોને નિયંત્રિત કરે છે જે જનીનો સાથે સંબંધ ધરાવતા નથી, તેના બદલે, તેઓ તંદુરસ્ત અને રોગવિજ્ઞાનવિષયક પરિસ્થિતિઓ હેઠળ કોષો, અવયવો અને આખરે સજીવો દ્વારા સામનો કરવામાં આવતા વિવિધ તણાવ-સંબંધિત સંજોગો સામે રક્ષણ આપે છે. પ્રકાશિત શૈક્ષણિક સંશોધન અભ્યાસોમાંથી માહિતીના આ નવા જથ્થાના આધારે, સંશોધકો હવે વધુ સારી રીતે વિકાસ કરી શકે છે Nrf2 આહાર પૂરવણીઓ.
2007 સુધીમાં, સંશોધન અભ્યાસોએ Nrf2 પાથવેના જટિલ કાર્યને દર્શાવ્યું છે. Nrf2 એક્ટિવેટર્સ માનવ શરીરની અંદર વિવિધ બંધારણોના પરિબળોની નકલ કરતા જોવા મળ્યા છે. આ માર્ગો દ્વારા, Nrf2 એક્ટિવેટર્સ સંતુલન જાળવવા અને જનીનોની વિકસતી આવશ્યકતાઓને પ્રતિસાદ આપવા માટે સમગ્ર કોષમાં બદલાતી પરિસ્થિતિઓને અનુભવવા માટે સજ્જ કરવામાં આવ્યા છે.
શા માટે Nrf2-એક્ટિવેટીંગ સપ્લિમેન્ટ્સનો ઉપયોગ કરવો?
સજીવોમાં વય સાથે Nrf2-સક્રિયકરણ ક્ષમતાઓ ઘટતી જાય છે, ફેરફારો થવાનું શરૂ થઈ શકે છે. સંશોધન અભ્યાસોએ દર્શાવ્યું છે કે કોષોમાં Nrf2 નું ધ્યાન વય સાથે ઘટતું જાય છે, જે ઓક્સિડેટીવ તણાવના વધતા માર્કર દર્શાવે છે. એથરોસ્ક્લેરોસિસ અને કાર્ડિયોવેસ્ક્યુલર રોગ, સંધિવા, કેન્સર, સ્થૂળતા, પ્રકાર 2 ડાયાબિટીસ, હાયપરટેન્શન, મોતિયા અને અલ્ઝાઈમર રોગ તેમજ પાર્કિન્સન રોગો જેવા વિવિધ વય-સંબંધિત રોગો આ ફેરફારોને કારણે વિકસી શકે છે. આ સ્વાસ્થ્ય સમસ્યાઓ સાથે ઓક્સિડેટીવ તણાવ જોવા મળ્યો છે.
Nrf2 એક્ટિવેટર્સનું ઉત્પાદન વધારવા માટે સેલની ક્ષમતાને ઉત્તેજીત કરીને, Nrf2 આહાર પૂરવણીઓ ઓક્સિડેટીવ તણાવની અસરોનો સામનો કરવા માટે માનવ શરીરની પોતાની ક્ષમતાને પુનર્જીવિત કરવામાં મદદ કરી શકે છે. પોલીઅનસેચ્યુરેટેડ ફેટી એસિડ્સ, અથવા PUFA, સૌથી સહેલાઈથી ઓક્સિડાઇઝ્ડ પરમાણુઓમાંના એક છે અને તેઓ ખાસ કરીને મુક્ત રેડિકલથી નુકસાન સહન કરવા માટે સંવેદનશીલ હોય છે. થિઓબાર્બિટ્યુરિક એસિડ, અથવા ટીબીએઆરએસ, ઉત્પાદન વય સાથે વધી શકે છે, જે એનઆરએફ2-નિયમિત માર્ગોમાં ઘટાડા સાથે ઓક્સિડેટીવ તણાવમાં વધારો સૂચવે છે.
જૈવિક રીતે, જનીન ઇન્ડક્શન એ ખરેખર ધીમી પદ્ધતિ છે, જેને સામાન્ય રીતે પાથવે દ્વારા સ્થાનાંતરિત કરવા માટે કલાકોની જરૂર પડે છે. પરિણામે, ઘણા ઉત્સેચકો પાસે તેમની પોતાની ઓન/ઓફ સ્વીચો હોય છે જે વિવિધ નિયમનકારી ઉત્સેચકો દ્વારા મિનિટોમાં ટ્રિગર થઈ શકે છે. સંશોધકોએ Nrf2 એક્ટિવેટર્સની માલિકીની રચનાઓ વિકસાવી છે જે સક્રિયકરણના આ જ્ઞાન આધારનો ઉપયોગ કરે છે. Nrf2 સક્રિયકરણ માત્ર Nrf2 ટ્રાન્સક્રિપ્શન પરિબળને તેના અવરોધકમાંથી છૂટા કરીને સેલ ન્યુક્લિયસમાં સ્થાનાંતરિત થતું નથી, પણ સાયટોપ્રોટેક્ટીવ જનીન અભિવ્યક્તિને પ્રોત્સાહિત કરવા માટે ચોક્કસ DNA ક્રમ સાથે બંધનકર્તા છે, જે Nrf2 ને ન્યુક્લિયસમાંથી બહાર કાઢવામાં આવે છે તેની ગતિને નિયંત્રિત કરે છે.
માનવ શરીરમાં Nrf2 ના સક્રિયકરણ અને નાબૂદીની પ્રક્રિયાને સમજવાથી સંશોધકોને તેના મોડ્યુલેશન દ્વારા જનીનોના પ્રતિબિંબને પરિપૂર્ણ કરવા માટે વિવિધ Nrf2 એક્ટિવેટર્સના સંયોજનો બનાવવાની મંજૂરી મળી છે. જ્ઞાન આધારના સંયોજન, અન્ય સંશોધન અભ્યાસોની વિશાળ વિવિધતા સાથે મળીને આહાર પૂરવણીઓ તરીકે ઉપયોગ માટે Nrf2 એક્ટિવેટર્સનું ઉત્પાદન કરવામાં મદદ કરી છે. અમારી માહિતીનો અવકાશ શિરોપ્રેક્ટિક અને કરોડરજ્જુના સ્વાસ્થ્ય સમસ્યાઓ સુધી મર્યાદિત છે. વિષયની ચર્ચા કરવા માટે, કૃપા કરીને ડૉ. જીમેનેઝને પૂછો અથવા અમારો સંપર્ક કરો915-850-0900 .
ઘૂંટણની પીડા એ જાણીતું લક્ષણ છે જે ઘૂંટણની વિવિધ ઇજાઓ અને/અથવા પરિસ્થિતિઓને કારણે થઈ શકે છે, જેમાં�રમતો ઇજાઓ. ઘૂંટણ એ માનવ શરીરના સૌથી જટિલ સાંધાઓમાંનું એક છે કારણ કે તે ચાર હાડકાં, ચાર અસ્થિબંધન, વિવિધ રજ્જૂ, બે મેનિસ્કી અને કોમલાસ્થિના આંતરછેદથી બનેલું છે. અમેરિકન એકેડેમી ઓફ ફેમિલી ફિઝિશિયન્સ અનુસાર, ઘૂંટણની પીડાના સૌથી સામાન્ય કારણોમાં પેટેલર સબલક્સેશન, પેટેલર ટેન્ડિનિટિસ અથવા જમ્પર્સ ઘૂંટણ અને ઓસ્ગુડ-સ્લેટર રોગનો સમાવેશ થાય છે. જોકે ઘૂંટણનો દુખાવો 60 વર્ષથી વધુ ઉંમરના લોકોમાં થવાની સંભાવના છે, પરંતુ ઘૂંટણનો દુખાવો બાળકો અને કિશોરોમાં પણ થઈ શકે છે. ઘૂંટણના દુખાવાની સારવાર RICE પદ્ધતિઓને અનુસરીને ઘરે કરી શકાય છે, જો કે, ઘૂંટણની ગંભીર ઇજાઓને તાત્કાલિક તબીબી સારવારની જરૂર પડી શકે છે, જેમાં શિરોપ્રેક્ટિક સંભાળનો સમાવેશ થાય છે.
એન્ટીઑકિસડન્ટોને વૈજ્ઞાનિક રીતે સંયોજનો તરીકે ઓળખવામાં આવે છે જે માનવ શરીરમાં ઓક્સિડેશન પ્રક્રિયાને પ્રતિબંધિત કરે છે, જેને જો અનચેક કરવામાં આવે તો, તે મુક્ત રેડિકલ બનાવી શકે છે જે અસંખ્ય સાંકળ પ્રતિક્રિયાઓ વિકસાવી શકે છે જે સેલ્યુલર નુકસાનનું કારણ બની શકે છે. સદભાગ્યે, માનવ શરીર આવા બિલ્ટ-ઇન રોગપ્રતિકારક તંત્ર બનાવી શકે છે, જો કે, જ્યારે માઉન્ટિંગ રિએક્ટિવ ઓક્સિજન પ્રજાતિઓ, અથવા આરઓએસ, તટસ્થ થવામાં અસમર્થ હોય, ત્યારે એક નાની જ્યોતની કલ્પના કરો જે ઓક્સિજન સાથે ભળી જાય ત્યારે નિયંત્રણમાંથી બહાર નીકળી જાય છે, નુકસાન થવાનું બંધાયેલ છે. .
જ્યોતના રૂપક પર વિસ્તરણ કરવાનું ચાલુ રાખવા માટે, ROS અથવા પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓની અસરને તટસ્થ કરવાની ક્ષમતા ન હોવાનું અંતિમ ઉત્પાદન નુકસાન તેમજ બળતરા છે, બીજા શબ્દોમાં કહીએ તો, માનવ શરીર તદ્દન શાબ્દિક રીતે આગમાં છે. અદ્ભુત બાબત એ છે કે ત્યાં એન્ટીઑકિસડન્ટો છે જે આ સ્વાસ્થ્ય સમસ્યા સામે લડવામાં જબરદસ્ત મદદ કરી શકે છે અને આ એન્ટીઑકિસડન્ટ ગ્લુટાથિઓન છે. જો કે 1889 માં જોવા મળ્યું હતું, ગ્લુટાથિઓનની એન્ટીઑકિસડન્ટ અસર આધુનિક સંશોધન અભ્યાસોમાં સૌથી રસપ્રદ વિષયો પૈકીનો એક બની ગયો છે.
એન્ટીઑકિસડન્ટોના માસ્ટર: ગ્લુટાથિઓન
શક્તિશાળી પદાર્થ એ ટ્રિપેપ્ટાઇડ છે જે સિસ્ટીન, ગ્લુટામિક એસિડ અને ગ્લાયસીનમાંથી વિકસે છે. મુક્ત રેડિકલની રચના સામે માનવ શરીરનું રક્ષણ કરવાની તેની ક્ષમતાને કારણે, ગ્લુટાથિઓન આખરે તંદુરસ્ત રોગપ્રતિકારક શક્તિને પ્રોત્સાહન આપવામાં મદદ કરી શકે છે. પર આધારિત છે 2015 માં વૈજ્ઞાનિક અહેવાલો, તે નિર્ધારિત કરવામાં આવ્યું હતું કે ગ્લુટાથિઓનની પેરોક્સિરેડિન અને કેટાલેઝ સાથે સિનર્જિસ્ટિક રીતે કાર્ય કરવાની ક્ષમતા હાઇડ્રોજન પેરોક્સાઇડ સામે કોષોનું રક્ષણ કરવામાં મદદ કરે છે. આ સિનર્જિસ્ટિક ફોર્મ્યુલા પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓ અથવા આરઓએસ સામે કાર્ય કરે છે. Glutathione, peroxidredin અને catalase એ સેલ્યુલર હોમિયોસ્ટેસિસના વધારામાં આવશ્યક તત્વો છે, જે તંદુરસ્ત કોષો, પેશીઓ અને અવયવોની એકસાથે આવશ્યક પ્રક્રિયા છે.
વધુમાં, ગ્લુટાથિઓન લિમ્ફોસાઇટના કાર્યો પર તેની મહત્વપૂર્ણ અસરનો ઉપયોગ કરીને એકંદર રોગપ્રતિકારક તંત્રની રચના અને કાર્યને વધારે છે. અનુસાર ઇમ્યુનોકેમિસ્ટ્રી વિભાગ, માનવ શરીરમાં ગ્લુટાથિઓનનું યોગ્ય રીતે પૂરક સ્તર રોગપ્રતિકારક પ્રતિક્રિયાઓને મોટા પ્રમાણમાં વધારી શકે છે. ઉદાહરણ તરીકે, બે રેન્ડમાઇઝ્ડ પ્લેસબો-નિયંત્રિત ટ્રાયલ્સ દર્શાવે છે કે એન-એસિટિલ-સિસ્ટીન, અથવા એનએસી સાથે રોગપ્રતિકારક-સમાધાનવાળા દર્દીઓની ઉપચારાત્મક સારવાર, બંને કિસ્સાઓમાં, મોટાભાગની રોગપ્રતિકારક પ્રક્રિયાઓમાં નોંધપાત્ર વૃદ્ધિમાં પરિણમે છે જેમાં સંપૂર્ણ કાયાકલ્પનો સમાવેશ થાય છે. કુદરતી કિલર સેલ પ્રવૃત્તિ. N-acetyl-cysteine, અથવા NAC, ગ્લુટાથિઓનમાંથી સલ્ફરનો ઉપયોગ કરે છે અને તેને ઝેરી અણુઓ સાથે જોડે છે, જે પછી પાણીમાં દ્રાવ્ય બને છે અને માનવ શરીરમાં વિસર્જન થાય છે.
ગ્લુટાથિઓન પાસે લિપોઇક એસિડને પુનઃજીવિત કરવાની તેમજ વિટામિન સી અને ઇને રિસાઇકલ કરવાની ક્ષમતા પણ છે, જે મુક્ત રેડિકલને બેઅસર કરવા માટે ઇલેક્ટ્રોન મોકલીને સિસ્ટમની ચોક્કસ પ્રક્રિયાઓ શરૂ કરવા માટે જરૂરી છે. ના સંશોધન અભ્યાસના આધારે PLOS ONE, ગ્લુટાથિઓન ડાયાબિટીસ મેટિલસ, અથવા T2DM, અને માયકોબેક્ટેરિયમ ટ્યુબરક્યુલોસિસવાળા દર્દીઓને અસર કરે છે. સામાન્ય રીતે, નબળી રોગપ્રતિકારક શક્તિ ધરાવતી વ્યક્તિઓ M. tb, અથવા માયકોબેક્ટેરિયમ ટ્યુબરક્યુલોસિસ, રોગ અથવા ચેપના વધુ સંપર્કમાં જોવાની વૃત્તિ ધરાવે છે. તદુપરાંત, T2DM વગરના લોકો કરતા T2 ની ટાઈપ 2 ડાયાબિટીસ મેટિલસ અથવા T2DM વાળા વ્યક્તિઓ TB ની શક્યતા બે થી ત્રણ ગણી વધારે છે. સંશોધન અભ્યાસમાં એવું પણ સૂચવવામાં આવ્યું છે કે T2DM ધરાવતા દર્દીઓથી અલગ મેક્રોફેજમાં ગ્લુટાથિઓનનું સ્તર વધારવાથી M.Tb રોગ અથવા ચેપના નિયંત્રણમાં સુધારો થાય છે. આ પરિણામો દર્શાવે છે કે TXNUMXDM ધરાવતા દર્દીઓમાં ગ્લુટાથિઓનનું નીચું સ્તર એમ. ટીબી રોગ અથવા ચેપની ઉચ્ચ તકમાં ફાળો આપે છે. તદુપરાંત, પર નિર્ભર છે બ્રાઇટન અને સસેક્સ મેડિકલ સ્કૂલમાં ડીટ્રો ગેઝી, ઓક્સિડેટીવ તણાવ આખરે નબળી રોગપ્રતિકારક તંત્રની રચના અને કાર્યનું કારણ બની શકે છે.
સદનસીબે, ગ્લુટાથિઓન રોગપ્રતિકારક શક્તિને મજબૂત અને નિયંત્રિત કરવામાં આવશ્યક ભૂમિકા ભજવે છે. દાખલા તરીકે, ટી-લિમ્ફોસાઇટ પ્રસાર, પોલીમોર્ફોન્યુક્લિયર ન્યુટ્રોફિલ્સની ફેગોસિટીક પ્રવૃત્તિ અને ડેંડ્રિટિક સેલ ફંક્શન્સ સહિત રોગપ્રતિકારક તંત્રમાં જન્મજાત અને અનુકૂલનશીલ પ્રક્રિયાઓ માટે ગ્લુટાથિઓન આવશ્યક છે, જે મૂળભૂત હોઈ શકે છે કારણ કે આ એન્ટિજેન-પ્રસ્તુત કોષોથી બનેલા છે. . સેલ-મેડિટેડ ઇમ્યુનિટીમાં પ્રોટીન એન્ટિજેન્સનો સમાવેશ થાય છે જે શરૂઆતમાં મેક્રોફેજ અને ડેંડ્રિટિક કોશિકાઓના એન્ડોસાયટીક વેસિકલ્સમાં અધોગતિ કરવાનું શરૂ કરે છે, તેથી, એન્ટિજેન-વિશિષ્ટ ટી કોશિકાઓના પ્રસારને સક્રિય કરવા માટે નાના પેપ્ટાઇડ્સ સપાટી પર દર્શાવવામાં આવે છે. વધુમાં, ગ્લુટાથિઓન સાયટોકાઇન્સના નિર્માણમાં મદદ કરે છે, અને ડેંડ્રિટિક કોષો દ્વારા ઇન્ટરફેરોન-ગામાનું ઉત્પાદન જાળવવું જરૂરી છે, જે માયકોબેક્ટેરિયા સહિતના અંતઃકોશિક પેથોજેન્સ સામે રક્ષણ આપવા માટે મહત્વપૂર્ણ છે.
N-acetyl-cysteine, અથવા NAC, જેને વૈજ્ઞાનિક રીતે ગ્લુટાથિઓનના પુરોગામી તરીકે ઓળખવામાં આવે છે, તે પણ એક ખૂબ જ શક્તિશાળી સેલ્યુલર એન્ટીઑકિસડન્ટ છે જેનો ઉપયોગ ફ્રી રેડિકલ સ્કેવેન્જર એન્ટીઑકિસડન્ટ તરીકે થાય છે. માં તેની ભૂમિકા માટે સામાન્ય રીતે ઓળખાય છે એસિટામિનોફેન ઝેરીતાને અટકાવે છે, NAC, or�N-acetyl-cysteine, ઘણા સ્વાસ્થ્ય અને સુખાકારી લાભો ધરાવવા માટે દર્શાવવામાં આવ્યું છે. અનુસાર સેલ જર્નલ, NAC તંદુરસ્ત દાહક પ્રતિભાવને ટેકો આપવામાં મદદ કરે છે અને માનવ મુદત અને અકાળ મજૂરી પર હકારાત્મક અસર કરી શકે છે. સંશોધન અભ્યાસે તારણ કાઢ્યું છે કે અગાઉના અકાળ જન્મ અને બેક્ટેરિયલ યોનિનોસિસ ધરાવતી સ્ત્રીઓમાં, ગર્ભાવસ્થાના 0.6મા અઠવાડિયા પછી પ્રોજેસ્ટેરોન સાથે દરરોજ 16 ગ્રામ એનએસી મૌખિક રીતે લેવામાં આવે છે, જે અકાળ જન્મ પુનરાવૃત્તિ અને સુધારેલ નવજાત પરિણામો સામે રક્ષણ આપે છે. નિષ્કર્ષમાં, સ્નાયુ નિર્માણ પર NAC ની સકારાત્મક અસરો પણ શોધી કાઢવામાં આવી હતી. ત્રણ મિનિટના સતત સંકોચન પછી, 15 ટકા ઉન્નત આઉટપુટ જોવા મળ્યું, જે દર્શાવે છે કે કેવી રીતે NAC સ્નાયુઓના નિર્માણમાં સુધારો કરવામાં અને પ્રસૂતિ દરમિયાન એકંદર થાક ઘટાડવામાં મૂળભૂત ભૂમિકા ભજવે છે.
સંશોધકોએ એ પણ શોધ્યું કે NAC, or�N-acetyl-cysteine, જેઓ પોલિસિસ્ટિક અંડાશયના સિન્ડ્રોમ, અથવા PCOS ધરાવતા હોય તેમને લાભ કરી શકે છે. પીસીઓએસ, અથવા પોલિસિસ્ટિક અંડાશયના સિન્ડ્રોમ, એ એક સામાન્ય અંતઃસ્ત્રાવી ગ્રંથિ-સંબંધિત રોગ છે જે લગભગ 5 થી 10 ટકા પ્રજનન વયની સ્ત્રીઓને અસર કરે છે. આવા દર્દીઓમાં, મેટાબોલિક સિન્ડ્રોમનો અનુભવ થવાનું વધુ જોખમ રહેલું છે, જ્યાં NAC નો ઉપયોગ તંદુરસ્ત ઇન્સ્યુલિન સ્તર અને સંવેદનશીલતાને પુનઃસ્થાપિત કરવામાં મદદ કરે છે.
ડૉ. એલેક્સ જિમેનેઝની આંતરદૃષ્ટિ
એકંદર આરોગ્ય અને સુખાકારી હાંસલ કરવામાં અને જાળવવામાં તેની મૂળભૂત ભૂમિકાને કારણે ગ્લુટાથિઓનને "એન્ટિઓક્સિડન્ટ્સના માસ્ટર" તરીકે ઓળખવામાં આવે છે. જ્યારે માનવ શરીર પોતાનું ગ્લુટાથિઓન ઉત્પન્ન કરવામાં સક્ષમ છે, નબળા પોષણ, પ્રદૂષણ, ઝેર, દવાઓ અને/અથવા દવાઓનો વધુ પડતો ઉપયોગ, તાણ, આઘાત, વૃદ્ધાવસ્થા, રોગ અને કિરણોત્સર્ગ આ બધા ગ્લુટાથિઓનના કુદરતી સ્તરને ઘટાડી શકે છે. આ બદલામાં વ્યક્તિઓને ઓક્સિડેટીવ તણાવ, મુક્ત રેડિકલ, ચેપ અને કેન્સરથી સેલ નુકસાન માટે વધુ સંવેદનશીલ બનાવી શકે છે. તેથી ગ્લુટાથિઓન પૂરક માનવ શરીર પર જબરદસ્ત ફાયદાઓ કરી શકે છે. વૈકલ્પિક સારવાર વિકલ્પો સાથે, જેમ કે ચિરોપ્રેક્ટિક સંભાળ, ગ્લુટાથિઓન સ્તરો ફરી એકવાર સુખાકારીને સુધારવા માટે નિયંત્રિત કરી શકાય છે.
વધુમાં, હેલ્થકેર પ્રોફેશનલ્સે અન્ય વૈકલ્પિક સારવાર વિકલ્પો સાથે ગ્લુટાથિઓન સપ્લીમેન્ટેશનનો ઉપયોગ અમલમાં મૂકવાનું સૂચન કર્યું છે, જેમ કે ચિરોપ્રેક્ટિક કાળજી, એકંદર આરોગ્ય અને સુખાકારીને વધુ સુધારવા માટે. એન્ટિઓક્સિડન્ટ્સ મહત્તમ સુખાકારી જાળવવા તેમજ કોષને નુકસાન અથવા નુકસાન પહોંચાડતા મુક્ત રેડિકલની સાંકળ પ્રતિક્રિયાને રોકવા માટે મહત્વપૂર્ણ છે. ગ્લુટાથિઓન જેવા શક્તિશાળી એન્ટીઑકિસડન્ટો, જેમ કે અગાઉ ઉપર ઉલ્લેખ કર્યો છે, આખરે આ મુક્ત રેડિકલના વિકાસને નિયંત્રિત કરવામાં મદદ કરે છે અને તંદુરસ્ત રોગપ્રતિકારક તંત્ર પ્રતિભાવ પ્રદાન કરે છે. સંશોધન અભ્યાસમાં જાણવા મળ્યું છે કે ચિરોપ્રેક્ટિક કાળજી કુદરતી રીતે માનવ શરીરમાં એન્ટીઑકિસડન્ટોની પ્રવૃત્તિમાં વધારો કરીને આ પ્રક્રિયામાં આવશ્યક ભૂમિકા ભજવી શકે છે. ચિરોપ્રેક્ટિક સંભાળ એ એક સલામત અને અસરકારક સારવાર અભિગમ છે જે કરોડરજ્જુની ખોટી ગોઠવણો, અથવા સબલક્સેશનને સુધારવા માટે કરોડરજ્જુના ગોઠવણો અને મેન્યુઅલ મેનિપ્યુલેશનનો ઉપયોગ કરે છે, જેથી માનવ શરીર દવાઓ/દવાઓ અને/અથવા સર્જિકલ હસ્તક્ષેપના ઉપયોગ વિના કુદરતી રીતે પોતાને સાજા કરી શકે.
અંતે, એન્ટીઑકિસડન્ટો તેમના જૈવિક ગુણધર્મોને ઘણા સ્વાસ્થ્ય લાભો દ્વારા પ્રદર્શિત કરે છે, જે આપણા આધુનિક વિશ્વમાં તણાવ, રોગ અને પ્રદૂષણના વધતા જતા આક્રમણ સાથે હવે પહેલા કરતા વધુ જરૂરી છે, જે બધા કોષોને નુકસાન અને/અથવા નુકસાનમાં ફાળો આપે છે. . ગ્લુટાથિઓન અને તેના પુરોગામી, NAC, or�N-acetyl-cysteine, એન્ટીઑકિસડન્ટોના ક્ષેત્રમાં તેમની શક્તિશાળી સ્થિતિ દર્શાવવાનું ચાલુ રાખે છે. વૈકલ્પિક સારવાર વિકલ્પો સાથે, જેમ કે ચિરોપ્રેક્ટિક સંભાળ, લોકો આ શક્તિશાળી એન્ટીઑકિસડન્ટ દ્વારા ઓફર કરવામાં આવતા તમામ લાભોનો લાભ લઈ શકે છે. અમારી માહિતીનો અવકાશ શિરોપ્રેક્ટિક તેમજ કરોડરજ્જુની ઇજાઓ અને સ્થિતિઓ સુધી મર્યાદિત છે. વિષયની ચર્ચા કરવા માટે, કૃપા કરીને ડૉ. જીમેનેઝને પૂછો અથવા અમારો સંપર્ક કરો915-850-0900 .
ડૉ. એલેક્સ જિમેનેઝ દ્વારા ક્યુરેટેડ
વધારાના વિષયો: પીઠનો દુખાવો
પીઠનો દુખાવો વિકલાંગતા માટેના સૌથી પ્રચલિત કારણોમાંનું એક અને કામ પરના દિવસો ચૂકી ગયા છે. વાસ્તવમાં, પીઠના દુખાવાને ડૉક્ટરની ઑફિસની મુલાકાતો માટેનું બીજું સૌથી સામાન્ય કારણ માનવામાં આવે છે, જે ફક્ત ઉપલા-શ્વસનતંત્રના ચેપથી વધુ છે. લગભગ 80 ટકા વસ્તી તેમના સમગ્ર જીવનમાં ઓછામાં ઓછા એક વખત પીઠનો દુખાવો અનુભવશે. કરોડરજ્જુ એ હાડકાં, સાંધા, અસ્થિબંધન અને સ્નાયુઓ, અન્ય નરમ પેશીઓની વચ્ચે બનેલી જટિલ રચના છે. આને કારણે, ઇજાઓ અને/અથવા વિકટ પરિસ્થિતિ, જેમ કે હર્નિયેટ ડિસ્ક, આખરે પીઠના દુખાવાના લક્ષણો તરફ દોરી શકે છે. રમતગમતની ઇજાઓ અથવા ઓટોમોબાઇલ અકસ્માતની ઇજાઓ પીઠના દુખાવા માટેનું સૌથી વારંવારનું કારણ છે, જો કે, કેટલીકવાર સરળ હલનચલન પીડાદાયક પરિણામો લાવી શકે છે. સદનસીબે, વૈકલ્પિક સારવાર વિકલ્પો, જેમ કે ચિરોપ્રેક્ટિક સંભાળ, કરોડરજ્જુના ગોઠવણો અને મેન્યુઅલ મેનિપ્યુલેશન્સના ઉપયોગ દ્વારા પીઠનો દુખાવો ઓછો કરવામાં મદદ કરી શકે છે, આખરે પીડા રાહતમાં સુધારો કરે છે.
વિજ્ઞાન આધારિત શિરોપ્રેક્ટર ડૉ. એલેક્ઝાન્ડર જિમેનેઝ પર એક નજર નાખે છે ઓક્સિડેટીવ તણાવ, તે શું છે, તે કેવી રીતે શરીરને અસર કરે છે અને પરિસ્થિતિને ઉકેલવા માટે એન્ટીઑકિસડન્ટ સંરક્ષણ.
એબ્સ્ટ્રેક્ટ: પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓ (ROS) સામાન્ય સેલ્યુલર ચયાપચય અને પર્યાવરણીય પરિબળો, જેમ કે હવા પ્રદૂષકો અથવા સિગારેટના ધુમાડાના પરિણામે જીવંત જીવો દ્વારા ઉત્પન્ન થાય છે. આરઓએસ અત્યંત પ્રતિક્રિયાશીલ પરમાણુઓ છે અને કાર્બોહાઇડ્રેટ્સ, ન્યુક્લિક એસિડ, લિપિડ્સ અને પ્રોટીન જેવા કોષની રચનાને નુકસાન પહોંચાડી શકે છે અને તેમના કાર્યોમાં ફેરફાર કરી શકે છે. ઓક્સિડન્ટ્સની તરફેણમાં ઓક્સિડન્ટ્સ અને એન્ટીઑકિસડન્ટો વચ્ચેના સંતુલનમાં ફેરફારને ઓક્સિડેટીવ સ્ટ્રેસ કહેવામાં આવે છે. સેલ સધ્ધરતા, સક્રિયકરણ, પ્રસાર અને અંગ કાર્ય માટે ઘટાડવા અને ઓક્સિડાઇઝિંગ (રેડોક્સ) સ્થિતિનું નિયમન મહત્વપૂર્ણ છે. એરોબિક સજીવોમાં સંકલિત એન્ટીઑકિસડન્ટ સિસ્ટમ્સ હોય છે, જેમાં એન્ઝાઈમેટિક અને નોન-એન્જાઈમેટિક એન્ટીઑકિસડન્ટોનો સમાવેશ થાય છે જે સામાન્ય રીતે ROS ની હાનિકારક અસરોને અવરોધિત કરવામાં અસરકારક હોય છે. જો કે, રોગવિજ્ઞાનવિષયક પરિસ્થિતિઓમાં, એન્ટીઑકિસડન્ટ સિસ્ટમો ભરાઈ શકે છે. ઓક્સિડેટીવ તાણ કેન્સર, ન્યુરોલોજીકલ ડિસઓર્ડર, એથરોસ્ક્લેરોસિસ, હાયપરટેન્શન, ઇસ્કેમિયા/પરફ્યુઝન, ડાયાબિટીસ, તીવ્ર શ્વસન તકલીફ સિન્ડ્રોમ, આઇડિયોપેથિક પલ્મોનરી ફાઇબ્રોસિસ, ક્રોનિક અવરોધક પલ્મોનરી રોગ અને અસ્થમા સહિતની ઘણી પેથોલોજીકલ પરિસ્થિતિઓ અને રોગોમાં ફાળો આપે છે. આ સમીક્ષામાં, અમે સેલ્યુલર ઓક્સિડન્ટ અને એન્ટીઑકિસડન્ટ સિસ્ટમ્સનો સારાંશ આપીએ છીએ અને ઓક્સિડેટીવ તણાવની સેલ્યુલર અસરો અને મિકેનિઝમ્સની ચર્ચા કરીએ છીએ.
મુખ્ય શબ્દો: એન્ટીઑકિસડન્ટ, ઓક્સિડન્ટ, ઓક્સિડેટીવ તણાવ, પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓ, રેડોક્સ
(WAO જર્નલ 2012; 5:9�19)
પ્રતિક્રિયાશીલ ઓક્સિજન પ્રજાતિઓ (ROS) સામાન્ય સેલ્યુલર ચયાપચયના પરિણામે જીવંત જીવો દ્વારા ઉત્પન્ન થાય છે. ઓછીથી મધ્યમ સાંદ્રતામાં, તેઓ શારીરિક કોષ પ્રક્રિયાઓમાં કાર્ય કરે છે, પરંતુ ઉચ્ચ સાંદ્રતામાં, તેઓ કોષના ઘટકોમાં પ્રતિકૂળ ફેરફારો પેદા કરે છે, જેમ કે લિપિડ્સ, પ્રોટીન અને ડીએનએ. ઓક્સિડેટીવ તાણ તરીકે ઓળખવામાં આવે છે. ઓક્સિડેટીવ તણાવ કેન્સર, ન્યુરોલોજીકલ ડિસઓર્ડર, 1�6 એથરોસ્ક્લેરોસિસ, હાયપરટેન્શન, ઇસ્કેમિયા/પરફ્યુઝન, 7�10 ડાયાબિટીસ, તીવ્ર શ્વસન તકલીફ સિન્ડ્રોમ, આઇડિયોપેથિક પલ્મોનરી ફાઇબ્રોસિસ, ક્રોનિક અવરોધક રોગ સહિત અનેક રોગવિજ્ઞાનવિષયક પરિસ્થિતિઓમાં ફાળો આપે છે. ,11 અને અસ્થમા.14�15 એરોબિક સજીવોમાં સંકલિત એન્ટીઑકિસડન્ટ પ્રણાલીઓ હોય છે, જેમાં એન્ઝાઈમેટિક અને નોનએનઝાઈમેટિક એન્ટીઑકિસડન્ટોનો સમાવેશ થાય છે જે સામાન્ય રીતે ROS ની હાનિકારક અસરોને અવરોધિત કરવામાં અસરકારક હોય છે. જો કે, રોગવિજ્ઞાનવિષયક પરિસ્થિતિઓમાં, એન્ટીઑકિસડન્ટ સિસ્ટમો ભરાઈ શકે છે. આ સમીક્ષામાં, અમે સેલ્યુલર ઓક્સિડન્ટ અને એન્ટીઑકિસડન્ટ સિસ્ટમ્સ અને આરોગ્ય અને રોગની સ્થિતિમાં ઘટાડો અને ઓક્સિડાઇઝિંગ (રેડોક્સ) સ્થિતિના નિયમનનો સારાંશ આપીએ છીએ.
ઓક્સિડન્ટ્સ
આરઓએસના અંતર્જાત સ્ત્રોતો
સામાન્ય સેલ્યુલર મેટાબોલિઝમના પરિણામે મોલેક્યુલર ઓક્સિજનમાંથી આરઓએસ ઉત્પન્ન થાય છે. આરઓએસને 2 જૂથોમાં વિભાજિત કરી શકાય છે: મુક્ત રેડિકલ અને નોનરેડિકલ. પરમાણુઓ જેમાં એક અથવા વધુ જોડી વગરના ઇલેક્ટ્રોન હોય છે અને આ રીતે પરમાણુને પ્રતિક્રિયા આપે છે તેને મુક્ત રેડિકલ કહેવામાં આવે છે. જ્યારે 2 મુક્ત રેડિકલ તેમના અનપેયર્ડ ઇલેક્ટ્રોનને વહેંચે છે, ત્યારે બિન-આમૂલ સ્વરૂપો બનાવવામાં આવે છે. 3 મુખ્ય આરઓએસ જે શારીરિક મહત્વ ધરાવે છે તે સુપરઓક્સાઇડ આયન (O22.), હાઇડ્રોક્સિલ રેડિકલ (OH), અને હાઇડ્રોજન પેરોક્સાઇડ (H2O2) છે. ROS નો સારાંશ કોષ્ટક 1 માં આપવામાં આવ્યો છે.
મોલેક્યુલર ઓક્સિજનમાં 1 ઈલેક્ટ્રોન ઉમેરવાથી સુપરઓક્સાઈડ આયનોની રચના થાય છે. 22 આ પ્રક્રિયા નિકોટિન એડેનાઈન ડાયન્યુક્લિયોટાઈડ ફોસ્ફેટ [NAD(P)H] ઓક્સિડેઝ અથવા ઝેન્થાઈન ઓક્સિડેઝ અથવા મિટોકોન્ડ્રીયલ ઈલેક્ટ્રોન ટ્રાન્સપોર્ટ સિસ્ટમ દ્વારા મધ્યસ્થી કરવામાં આવે છે. સુપરઓક્સાઇડ આયનોના ઉત્પાદન માટેનું મુખ્ય સ્થળ મિટોકોન્ડ્રિયા છે, એડેનોસિન ટ્રાઇફોસ્ફેટ ઉત્પન્ન કરવા માટે કોષની મશીનરી. સામાન્ય રીતે, ઇલેક્ટ્રોનને પાણીમાં ઓક્સિજન ઘટાડવા માટે મિટોકોન્ડ્રીયલ ઇલેક્ટ્રોન પરિવહન સાંકળ દ્વારા સ્થાનાંતરિત કરવામાં આવે છે, પરંતુ લગભગ 1 થી 3% બધા ઇલેક્ટ્રોન સિસ્ટમમાંથી લીક થાય છે અને સુપરઓક્સાઇડ ઉત્પન્ન કરે છે. NAD(P)H ઓક્સિડેઝ પોલીમોર્ફોન્યુક્લિયર લ્યુકોસાઈટ્સ, મોનોસાઈટ્સ અને મેક્રોફેજમાં જોવા મળે છે. ફેગોસિટોસિસ પર, આ કોષો સુપરઓક્સાઇડનો વિસ્ફોટ ઉત્પન્ન કરે છે જે બેક્ટેરિયાનાશક પ્રવૃત્તિ તરફ દોરી જાય છે. સુપરઓક્સાઇડ ડિસમ્યુટેસિસ (SODs, EC 1.15.1.1) ની ક્રિયા દ્વારા સુપરઓક્સાઇડ હાઇડ્રોજન પેરોક્સાઇડમાં રૂપાંતરિત થાય છે. હાઇડ્રોજન પેરોક્સાઇડ સરળતાથી પ્લાઝ્મા પટલમાં ફેલાય છે. હાઇડ્રોજન પેરોક્સાઇડ પણ ઝેન્થાઇન ઓક્સિડેઝ, એમિનો એસિડ ઓક્સિડેઝ અને NAD(P)H ઓક્સિડેઝ23,24 અને પેરોક્સિસોમમાં ચયાપચયની પ્રતિક્રિયાઓમાં મોલેક્યુલર ઓક્સિજનના વપરાશ દ્વારા ઉત્પન્ન થાય છે. હેબર વેઈસ અને ફેન્ટન પ્રતિક્રિયાઓ નામની પ્રતિક્રિયાઓના અનુગામી સ્વરૂપમાં, H2O2 ફે2 અથવા Cu21 જેવી ટ્રાન્સમિશન ધાતુઓની હાજરીમાં OH21.25 માં ભંગાણ કરી શકે છે.
Fe31 +�.O2�?Fe2 +�O2 હેબર વેઇસ
Fe2 +�H2O2�?Fe3 +�OH�+ .OH ફેન્ટન પ્રતિક્રિયા
O 2 પોતે H2 O2 સાથે પણ પ્રતિક્રિયા કરી શકે છે અને OH�.26,27 હાઇડ્રોક્સિલ રેડિકલ ROS નું સૌથી વધુ પ્રતિક્રિયાશીલ છે અને પ્રોટીન, લિપિડ અને કાર્બોહાઇડ્રેટ્સ અને DNA ને નુકસાન પહોંચાડી શકે છે. તે બહુઅસંતૃપ્ત ફેટી એસિડ્સમાંથી ઇલેક્ટ્રોન લઈને લિપિડ પેરોક્સિડેશન પણ શરૂ કરી શકે છે.
ગ્રાન્યુલોસાયટીક ઉત્સેચકો ઇઓસિનોફિલ પેરોક્સિડેઝ અને માયલોપેરોક્સિડેઝ (MPO) દ્વારા H2O2 ની પ્રતિક્રિયાશીલતાને વધુ વિસ્તૃત કરે છે. સક્રિય ન્યુટ્રોફિલ્સમાં, H2O2 એમપીઓ દ્વારા લેવામાં આવે છે. ક્લોરાઇડ આયનની હાજરીમાં, H2O2 હાયપોક્લોરસ એસિડ (HOCl) માં રૂપાંતરિત થાય છે. HOCl અત્યંત ઓક્સિડેટીવ છે અને વાયુમાર્ગમાં પેથોજેન્સને મારી નાખવામાં મહત્વની ભૂમિકા ભજવે છે. 28 જો કે, HOCl ડીએનએ સાથે પ્રતિક્રિયા પણ કરી શકે છે અને ડીએનએ-પ્રોટીન ક્રિયાપ્રતિક્રિયાઓને પ્રેરિત કરી શકે છે અને પાયરીમિડીન ઓક્સિડેશન ઉત્પાદનો ઉત્પન્ન કરી શકે છે અને ડીએનએ બેઝમાં ક્લોરાઇડ ઉમેરી શકે છે. 29,30 ઇઓસિનોફિલ પેરોક્સિડેઝ અને એમપીઓ ટાયરોસિલ રેડિકલ દ્વારા હેલોજનેશન, નાઈટ્રેશન અને પ્રોટીન ક્રોસ-લિંક દ્વારા પ્રોટીનમાં ફેરફાર કરીને ઓક્સિડેટીવ તણાવમાં પણ ફાળો આપે છે.31�33
અન્ય ઓક્સિજન-પ્રાપ્ત મુક્ત રેડિકલ પેરોક્સિલ રેડિકલ (ROO$) છે. આ રેડિકલનું સૌથી સરળ સ્વરૂપ હાઇડ્રો-પેરોક્સિલ રેડિકલ (HOO$) છે અને ફેટી એસિડ પેરોક્સિડેશનમાં તેની ભૂમિકા છે. મુક્ત રેડિકલ સાઇડ-ચેઇન મેથિલિન કાર્બનમાંથી હાઇડ્રોજન અણુને અમૂર્ત કરીને લિપિડ પેરોક્સિડેશન સાંકળ પ્રતિક્રિયાઓને ઉત્તેજિત કરી શકે છે. લિપિડ રેડિકલ પછી ઓક્સિજન સાથે પ્રતિક્રિયા કરીને પેરોક્સિલ રેડિકલ ઉત્પન્ન કરે છે. પેરોક્સિલ રેડિકલ સાંકળ પ્રતિક્રિયા શરૂ કરે છે અને બહુઅસંતૃપ્ત ફેટી એસિડને લિપિડ હાઇડ્રોપેરોક્સાઇડમાં પરિવર્તિત કરે છે. લિપિડ હાઇડ્રોપેરોક્સાઇડ્સ ખૂબ જ અસ્થિર છે અને એલ્ડીહાઇડ્સ (જેમ કે 4-હાઇડ્રોક્સી-2,3-નોનેનલ) અને મેલોન્ડિઆલ્ડીહાઇડ્સ (એમડીએ) જેવા ગૌણ ઉત્પાદનોમાં સરળતાથી વિઘટિત થાય છે. આઇસોપ્રોસ્ટેનેસ એ લિપિડ પેરોક્સિડેશન ઉત્પાદનોનું બીજું જૂથ છે જે એરાકીડોનિક એસિડના પેરોક્સિડેશન દ્વારા ઉત્પન્ન થાય છે અને તે અસ્થમાના પ્લાઝ્મા અને શ્વાસના ઘનીકરણમાં પણ ઉન્નત હોવાનું જણાયું છે. 34,35 લિપિડનું પેરોક્સિડેશન કોષ પટલની અખંડિતતાને ખલેલ પહોંચાડે છે અને પુનઃપ્રાપ્તિ તરફ દોરી જાય છે. પટલ માળખું.
હાઇડ્રોજન પેરોક્સાઇડ, સુપરઓક્સાઇડ રેડિકલ, ઓક્સિડાઇઝ્ડ ગ્લુટાથિઓન (GSSG), MDAs, isoprostanes, carbonyls, અને nitrotyrosine ને પ્લાઝ્મા, રક્ત અથવા બ્રોન્કોઆલ્વીઓલર લેવેજ નમૂનાઓમાંથી પ્રમાણિત તપાસ દ્વારા ઓક્સિડેશનના બાયોમાર્કર્સ તરીકે સરળતાથી માપી શકાય છે.
ઓક્સિડન્ટ્સનો બાહ્ય સ્ત્રોત
સિગારેટનો ધુમાડો
સિગારેટના ધુમાડામાં ઘણા ઓક્સિડન્ટ્સ અને ફ્રી રેડિકલ અને કાર્બનિક સંયોજનો હોય છે, જેમ કે સુપરઓક્સાઇડ અને નાઈટ્રિક ઑક્સાઈડ. 36 વધુમાં, સિગારેટના ધુમાડાને ફેફસામાં શ્વાસમાં લેવાથી કેટલાક અંતર્જાત મિકેનિઝમ્સ પણ સક્રિય થાય છે, જેમ કે ન્યુટ્રોફિલ્સ અને મેક્રોફેજનું સંચય, જે ઓક્સિડેન્ટમાં વધુ વધારો કરે છે. .
ઓઝોન એક્સપોઝર
ઓઝોન એક્સપોઝર લિપિડ પેરોક્સિડેશનનું કારણ બની શકે છે અને વાયુમાર્ગ ઉપકલામાં ન્યુટ્રોફિલ્સના પ્રવાહને પ્રેરિત કરી શકે છે. ઓઝોનના ટૂંકા ગાળાના સંસર્ગમાં એમપીઓ, ઇઓસિનોફિલ કેશનીક પ્રોટીન અને લેક્ટેટ ડીહાઇડ્રોજેનેઝ અને આલ્બ્યુમિન જેવા બળતરા મધ્યસ્થીઓના પ્રકાશનનું કારણ પણ બને છે. 37 તંદુરસ્ત વિષયોમાં પણ, ઓઝોનના સંપર્કમાં પલ્મોનરી કાર્યોમાં ઘટાડો થાય છે. 38 Cho et al39 દર્શાવે છે કે સૂક્ષ્મ દ્રવ્ય (ઘન કણો અને પ્રવાહી ટીપાંનું મિશ્રણ હવામાં અટકી જાય છે) ઓક્સિજનના ઘટાડાને ઉત્પ્રેરિત કરે છે.
હાયપરૉક્સિયા
હાયપરૉક્સિયા ફેફસાં અથવા શરીરના અન્ય પેશીઓમાં ઑક્સિજનના સામાન્ય આંશિક દબાણ કરતાં ઊંચા ઓક્સિજન સ્તરની સ્થિતિનો સંદર્ભ આપે છે. તે પ્રતિક્રિયાશીલ ઓક્સિજન અને નાઇટ્રોજન પ્રજાતિઓના વધુ ઉત્પાદન તરફ દોરી જાય છે. 40,41
આયોનાઇઝિંગ રેડિએશન
આયોનાઇઝિંગ રેડિયેશન, O2 ની હાજરીમાં, હાઇડ્રોક્સિલ રેડિકલ, સુપરઓક્સાઇડ અને ઓર્ગેનિક રેડિકલને હાઇડ્રોજન પેરોક્સાઇડ અને ઓર્ગેનિક હાઇડ્રોપેરોક્સાઇડમાં રૂપાંતરિત કરે છે. આ હાઇડ્રોપેરોક્સાઇડ પ્રજાતિઓ ફેન્ટન પ્રતિક્રિયાઓ દ્વારા ફે અને ક્યુ જેવા રેડોક્સ સક્રિય ધાતુના આયનો સાથે પ્રતિક્રિયા આપે છે અને આમ ઓક્સિડેટીવ તણાવને પ્રેરિત કરે છે. 42,43 નારાયણન એટ અલ 44 દર્શાવે છે કે આલ્ફા કણોના સંપર્કમાં આવતા ફાઇબ્રોબ્લાસ્ટ્સ અંતઃકોશિક O2 2 અને H2O2 માં નોંધપાત્ર વધારો કરે છે. પ્લાઝ્મા મેમ્બ્રેન-બાઉન્ડ NADPH ઓક્સિડેઝ દ્વારા ઉત્પાદન.44 સિગ્નલ ટ્રાન્સડક્શન પરમાણુઓ, જેમ કે એક્સ્ટ્રા સેલ્યુલર સિગ્નલ-રેગ્યુલેટેડ કિનેઝ 1 અને 2 (ERK1/2), c-જુન એન-ટર્મિનલ કિનેઝ (JNK), અને p38, અને ટ્રાન્સક્રિપ્શન પરિબળો, જેમ કે એક્ટિવેટર પ્રોટીન-1 (AP-1), ન્યુક્લિયર ફેક્ટર-kB (NF-kB), અને p53, સક્રિય થાય છે, જેના પરિણામે રેડિયેશન રિસ્પોન્સ સંબંધિત જીન્સની અભિવ્યક્તિ થાય છે. પોર્ફિરિન્સ, NADPH ઓક્સિડેઝ અને રિબોફ્લેવિન્સ જેવા અંતર્જાત ફોટોસેન્સિટાઇઝર્સના ઉત્તેજના દ્વારા. 45-Oxo-50- dihydroguanine (8-oxoGua) મુખ્ય UVA- મધ્યસ્થી DNA ઓક્સિડેશન ઉત્પાદન છે જે OH રેડિકલ, 7,8-ઇલેક્ટ્રોન ઓક્સિડન્ટ્સ અને સિંગલ ઓક્સિજનના ઓક્સિડેશન દ્વારા રચાય છે જે મુખ્યત્વે ગ્વાનિન સાથે પ્રતિક્રિયા આપે છે. 8 ગ્વાનિનની રચના આઇસોલેટેડ ડીએનએમાં રેડિકલ કેશન આયનાઇઝિંગ રેડિયેશનની સીધી અસર દ્વારા અસરકારક રીતે જોવા મળે છે. 1 આયનાઇઝિંગ રેડિયેશનના સંપર્કમાં આવ્યા પછી, ગ્લુટાથિઓન (GSH) નું ઇન્ટ્રાસેલ્યુલર સ્તર ટૂંકા ગાળા માટે ઘટે છે પરંતુ તે પછી ફરી વધે છે.51
હેવી મેટલ આયનો
ભારે ધાતુના આયનો, જેમ કે આયર્ન, કોપર, કેડમિયમ, પારો, નિકલ, સીસું અને આર્સેનિક, પ્રતિક્રિયાશીલ રેડિકલના ઉત્પાદનને પ્રેરિત કરી શકે છે અને લિપિડ પેરોક્સિડેશન અને અણુ પ્રોટીન અને ડીએનએ સાથે પ્રતિક્રિયા દ્વારા એન્ઝાઇમ પ્રવૃત્તિઓના અવક્ષય દ્વારા સેલ્યુલર નુકસાનનું કારણ બની શકે છે.
મેટલ-મધ્યસ્થી મુક્ત રેડિકલ જનરેશનની સૌથી મહત્વપૂર્ણ પદ્ધતિઓ પૈકીની એક ફેન્ટન-પ્રકારની પ્રતિક્રિયા છે. સુપરઓક્સાઇડ આયન અને હાઇડ્રોજન પેરોક્સાઇડ સંક્રમણ ધાતુઓ સાથે ક્રિયાપ્રતિક્રિયા કરી શકે છે, જેમ કે આયર્ન અને કોપર, ધાતુના ઉત્પ્રેરિત હેબર વેઇસ/ફેન્ટન પ્રતિક્રિયા દ્વારા OH રેડિકલ બનાવવા માટે.
ફેન્ટન-પ્રકાર અને હેબર-વેઇસ-પ્રકારની પદ્ધતિઓ ઉપરાંત, અમુક ધાતુના આયનો સેલ્યુલર પરમાણુઓ સાથે સીધી પ્રતિક્રિયા આપી શકે છે જેથી મુક્ત રેડિકલ ઉત્પન્ન થાય, જેમ કે થિયોલ રેડિકલ, અથવા સેલ સિગ્નલિંગ પાથવેને પ્રેરિત કરે. આ રેડિકલ O22.. O22 પેદા કરવા માટે અન્ય થિયોલ પરમાણુઓ સાથે પણ પ્રતિક્રિયા આપી શકે છે. H2O2 માં રૂપાંતરિત થાય છે, જે વધારાના ઓક્સિજન રેડિકલ જનરેશનનું કારણ બને છે. કેટલીક ધાતુઓ, જેમ કે આર્સેનાઈટ, કોષોમાં આમૂલ ઉત્પાદન પ્રણાલીના સક્રિયકરણ દ્વારા પરોક્ષ રીતે આરઓએસ રચનાને પ્રેરિત કરે છે.56
આર્સેનિક એ અત્યંત ઝેરી તત્વ છે જે સુપરઓક્સાઈડ (O2 2), સિંગલ ઓક્સિજન (1O2), પેરોક્સાઈલ રેડિકલ (ROO), નાઈટ્રિક ઓક્સાઈડ (NO ), હાઈડ્રોજન પેરોક્સાઇડ (H2O2), અને ડાયમેથિલાર્સિનીક પેરોક્સાઈલ રેડિકલ [( CH3)2AsOO ].57�59 આર્સેનિક (III) સંયોજનો એન્ટીઑકિસડન્ટ ઉત્સેચકોને અટકાવી શકે છે, ખાસ કરીને GSH-આશ્રિત ઉત્સેચકો, જેમ કે glutathione-S-transferases (GSTs), glutathione peroxidase (GSH-Px), અને GSH રિડક્ટેઝ, બાઇન્ડ દ્વારા. - તેમના સલ્ફહાઇડ્રેલ (�SH) જૂથોમાં સામેલ છે.60,61
લીડ લિપિડ પેરોક્સિડેશનમાં વધારો કરે છે. 62 ટીશ્યુ SOD અને મગજ GPx ની પ્રવૃત્તિમાં નોંધપાત્ર ઘટાડો લીડના એક્સપોઝર પછી નોંધવામાં આવ્યો છે. 63,64 ઝીંકનું રિપ્લેસમેન્ટ, જે લીડ દ્વારા ઘણા ઉત્સેચકો માટે કોફેક્ટર તરીકે કામ કરે છે, આવા ઉત્સેચકોની નિષ્ક્રિયતા તરફ દોરી જાય છે. લીડ એક્સપોઝર ટીશ્યુ થિયોલ્સને અસર કરીને GST ના નિષેધનું કારણ બની શકે છે.
મેટલ-ઉત્પ્રેરિત પ્રતિક્રિયાઓ દ્વારા જનરેટ થયેલ આરઓએસ ડીએનએ પાયામાં ફેરફાર કરી શકે છે. ત્રણ બેઝ અવેજી, G/C, G/T, અને C/T, મેટલ આયનો દ્વારા ઓક્સિડેટીવ નુકસાનના પરિણામે થઈ શકે છે, જેમ કે Fe21, Cu21 અને Ni21. Reid et al65 એ દર્શાવ્યું હતું કે G/C મુખ્યત્વે Fe21 દ્વારા ઉત્પાદિત કરવામાં આવ્યું હતું જ્યારે C/T અવેજીકરણ Cu21 અને Ni21 દ્વારા કરવામાં આવ્યું હતું.
એન્ટિઓક્સિડન્ટ્સ
માનવ શરીર વિવિધ પ્રકારના એન્ટીઑકિસડન્ટોથી સજ્જ છે જે ઑક્સિડન્ટ્સની અસરને સંતુલિત કરવા માટે સેવા આપે છે. તમામ વ્યવહારુ હેતુઓ માટે, આને 2 શ્રેણીઓમાં વિભાજિત કરી શકાય છે: એન્ઝાઈમેટિક (કોષ્ટક 2) અને નોનએનઝાઈમેટિક (કોષ્ટક 3).
એન્ઝાઇમેટિક એન્ટીઑકિસડન્ટો
ફેફસાંના મુખ્ય એન્ઝાઇમેટિક એન્ટીઑકિસડન્ટો SODs (EC 1.15.1.11), catalase (EC 1.11.1.6), અને GSH-Px (EC 1.11.1.9) છે. આ મુખ્ય ઉત્સેચકો ઉપરાંત, અન્ય એન્ટીઑકિસડન્ટો, જેમાં હેમ ઓક્સિજેનેઝ-1 (EC 1.14.99.3), અને રેડોક્સ પ્રોટીન, જેમ કે થિયોરેડોક્સિન્સ (TRXs, EC 1.8.4.10), પેરોક્સિરેડોક્સિન્સ (PRXs, EC 1.11.1.15), અને glutaroxins. , પલ્મોનરી એન્ટીઑકિસડન્ટ સંરક્ષણમાં નિર્ણાયક ભૂમિકા ભજવતા હોવાનું પણ જાણવા મળ્યું છે.
સુપરઓક્સાઇડ એ વિવિધ સ્ત્રોતોમાંથી ઉત્પાદિત પ્રાથમિક આરઓએસ હોવાથી, દરેક કોષ માટે એસઓડી દ્વારા તેનું વિસર્જન પ્રાથમિક મહત્વ છે. SOD ના તમામ 3 સ્વરૂપો, એટલે કે, CuZn-SOD, Mn-SOD અને EC-SOD, માનવ ફેફસામાં વ્યાપકપણે વ્યક્ત થાય છે. Mn-SOD એ મિટોકોન્ડ્રિયા મેટ્રિક્સમાં સ્થાનીકૃત છે. EC-SOD મુખ્યત્વે એક્સ્ટ્રા સેલ્યુલર મેટ્રિક્સમાં સ્થાનીકૃત છે, ખાસ કરીને એવા વિસ્તારોમાં જેમાં ઉચ્ચ માત્રામાં પ્રકાર I કોલેજન ફાઇબર હોય છે અને પલ્મોનરી અને પ્રણાલીગત જહાજોની આસપાસ હોય છે. તે શ્વાસનળીના ઉપકલા, મૂર્ધન્ય ઉપકલા અને મૂર્ધન્ય મેક્રોફેજેસમાં પણ મળી આવ્યું છે. 66,67 એકંદરે, CuZn-SOD અને Mn-SOD સામાન્ય રીતે સુપરઓક્સાઇડ રેડિકલના બલ્ક સ્કેવેન્જર્સ તરીકે કામ કરે છે. ફેફસાંમાં પ્રમાણમાં ઊંચું EC-SOD સ્તર બાહ્યકોષીય મેટ્રિક્સ ઘટકો સાથે તેના ચોક્કસ બંધન સાથે ફેફસાના મેટ્રિક્સ સંરક્ષણના મૂળભૂત ઘટકનું પ્રતિનિધિત્વ કરી શકે છે.
H2O2 કે જે SOD ની ક્રિયા અથવા ઓક્સિડેઝની ક્રિયા દ્વારા ઉત્પન્ન થાય છે, જેમ કે xanthine oxidase, catalase અને GSH-Px દ્વારા પાણીમાં ઘટાડો થાય છે. કેટાલેઝ 4 સમાન મોનોમર્સથી બનેલા ટેટ્રામર તરીકે અસ્તિત્વમાં છે, જેમાંથી દરેક સક્રિય સાઇટ પર હેમ જૂથ ધરાવે છે. H2O2 નું અધોગતિ કેટાલેઝ-ફેરીકેટલેઝ (પાણી સાથે સંકલિત આયર્ન) અને સંયોજન I (ઓક્સિજન પરમાણુ સાથે સંકુલિત આયર્ન) વચ્ચેના રૂપાંતરણ દ્વારા પરિપૂર્ણ થાય છે. H2O2 દ્વારા એન્ઝાઇમના ઓક્સિડેટીવ નિષ્ક્રિયકરણ (કમ્પાઉન્ડ II ની રચના) અટકાવવા માટે કેટાલેઝ એનએડીપીએચને ઘટાડતા સમકક્ષ તરીકે પણ બાંધે છે કારણ કે તે પાણીમાં ઘટાડી દેવામાં આવે છે.2
H2O2 અને લિપિડ હાઇડ્રોપેરોક્સાઇડના ઘટાડા માટે જવાબદાર રેડોક્સ ચક્રના ઉત્સેચકોમાં GSH-Pxs નો સમાવેશ થાય છે. 70 GSH-Pxs એ ટેટ્રામેરિક ઉત્સેચકોનું એક કુટુંબ છે જેમાં વિશિષ્ટ એમિનો એસિડ સેલેનોસિસ્ટીન હોય છે. સક્રિય સાઇટ્સ અને H2O2 અને લિપિડ પેરોક્સાઇડ્સને તેમના અનુરૂપ આલ્કોહોલમાં ઘટાડવા માટે ઓછા પરમાણુ-વજનના થિયોલ્સનો ઉપયોગ કરે છે, જેમ કે GSH. ચાર GSH- Pxs વર્ણવવામાં આવ્યા છે, જે વિવિધ જનીનો દ્વારા એન્કોડ કરવામાં આવ્યા છે: GSH- Px-1 (સેલ્યુલર GSH-Px) સર્વવ્યાપક છે અને H2O2 અને ફેટી એસિડ પેરોક્સાઇડ્સ ઘટાડે છે, પરંતુ એસ્ટરિફાઈડ પેરોક્સાઈલ લિપિડ્સ નથી. 71 એસ્ટરિફાઈડ લિપિડ્સ મેમ્બ્રેન-બાઉન્ડ GSH દ્વારા ઘટાડવામાં આવે છે. -Px-4 (ફોસ્ફોલિપીડ હાઇડ્રોપેરોક્સાઇડ GSH-Px), જે સમકક્ષ ઘટાડવા માટે ઘણા જુદા-જુદા ઓછા-મોલેક્યુલર-વજન થિયોલ્સનો ઉપયોગ કરી શકે છે. GSH-Px-2 (ગેસ્ટ્રોઇન્ટેસ્ટાઇનલ GSH-Px) જઠરાંત્રિય ઉપકલા કોષોમાં સ્થાનીકૃત છે જ્યાં તે આહારના પેરોક્સાઇડ્સને ઘટાડવા માટે સેવા આપે છે. એક્સ્ટ્રા સેલ્યુલર કમ્પાર્ટમેન્ટ અને સસ્તન પ્રાણીઓમાં સૌથી મહત્વપૂર્ણ એક્સ્ટ્રા સેલ્યુલર એન્ટીઑકિસડન્ટ એન્ઝાઇમ માનવામાં આવે છે. આમાંથી, બાહ્યકોષીય GSH-Px માનવ ફેફસામાં સૌથી વધુ વ્યાપકપણે તપાસવામાં આવે છે.72
વધુમાં, H2O2 નો નિકાલ ઘણા થિયોલ-સમાવતી ઉત્સેચકો સાથે ગાઢ રીતે સંકળાયેલો છે, જેમ કે, TRXs (TRX1 અને TRX2), થિયોરેડોક્સિન રિડક્ટેસિસ (EC 1.8.1.9) (TRRs), PRXs (જે થિયોરેડોક્સિન પેરોક્સિડેસિસ છે), અને ગ્લુટારેડોક્સિન 74.
બે TRXs અને TRRs માનવ કોષોમાં લાક્ષણિકતા ધરાવે છે, જે સાયટોસોલ અને મિટોકોન્ડ્રિયા બંનેમાં અસ્તિત્વ ધરાવે છે. ફેફસામાં, TRX અને TRR શ્વાસનળી અને મૂર્ધન્ય ઉપકલા અને મેક્રોફેજેસમાં વ્યક્ત થાય છે. માનવ કોષોમાં છ અલગ-અલગ PRX મળી આવ્યા છે, જે તેમના અલ્ટ્રાસ્ટ્રક્ચરલ કમ્પાર્ટમેન્ટલાઇઝેશનમાં અલગ છે. પ્રાયોગિક અભ્યાસોએ મૂર્ધન્ય ઉપકલાના રક્ષણમાં PRX VI નું મહત્વ જાહેર કર્યું છે. માનવ ફેફસાં શ્વાસનળીના ઉપકલા, મૂર્ધન્ય ઉપકલા અને મેક્રોફેજેસમાં તમામ PRX ને વ્યક્ત કરે છે. 75 PRX V તાજેતરમાં પેરોક્સિનાઇટ્રેટ રિડક્ટેઝ તરીકે કામ કરતું જણાયું છે,76 જેનો અર્થ છે કે તે ROS-મધ્યસ્થી ફેફસાની ઇજાના વિકાસમાં સંભવિત રક્ષણાત્મક સંયોજન તરીકે કાર્ય કરી શકે છે. .77
આ એન્ટીઑકિસડન્ટો માટે સામાન્ય છે NADPH ની જરૂરિયાત ઘટાડતા સમકક્ષ તરીકે. NADPH સક્રિય સ્વરૂપમાં કેટાલેઝ જાળવી રાખે છે અને તેનો ઉપયોગ TRX અને GSH રિડક્ટેઝ (EC 1.6.4.2) દ્વારા કોફેક્ટર તરીકે થાય છે, જે GSSG ને GSH માં રૂપાંતરિત કરે છે, જે GSH-Pxs માટે સહ-સબસ્ટ્રેટ છે. ઇન્ટ્રાસેલ્યુલર NADPH, બદલામાં, ગ્લુકોઝ-1-ફોસ્ફેટ ડિહાઇડ્રોજેનેઝ દ્વારા NADP6 ના ઘટાડા દ્વારા ઉત્પન્ન થાય છે, જે પેન્ટોઝ ફોસ્ફેટ પાથવેના પ્રથમ અને દર-મર્યાદિત એન્ઝાઇમ છે, જે ગ્લુકોઝ-6-ફોસ્ફેટના 6-ફોસ્ફેક્ટોન્ગ્લૂન રૂપાંતર દરમિયાન થાય છે. NADPH જનરેટ કરીને, ગ્લુકોઝ-6-ફોસ્ફેટ ડિહાઈડ્રોજેનેઝ એ સાયટોસોલિક GSH બફરિંગ ક્ષમતા (GSH/ GSSG) નું નિર્ણાયક નિર્ણાયક છે અને તેથી, એક આવશ્યક, નિયમનકારી એન્ટીઑકિસડન્ટ એન્ઝાઇમ ગણી શકાય. 78,79
GSTs (EC 2.5.1.18), અન્ય એન્ટીઑકિસડન્ટ એન્ઝાઇમ કુટુંબ, ગૌણ ચયાપચયને નિષ્ક્રિય કરે છે, જેમ કે અસંતૃપ્ત એલ્ડીહાઇડ્સ, ઇપોક્સાઇડ્સ અને હાઇડ્રોપેરોક્સાઇડ્સ. GST ના ત્રણ મુખ્ય પરિવારોનું વર્ણન કરવામાં આવ્યું છે: સાયટોસોલિક GST, માઇટોકોન્ડ્રીયલ GST,80,81 અને મેમ્બ્રેન-સંબંધિત માઇક્રોસોમલ GST જે ઇકોસોનોઇડ અને GSH ચયાપચયમાં ભૂમિકા ધરાવે છે. 82 સાયટોસોલિક GST ના સાત વર્ગો સસ્તન પ્રાણીઓમાં ઓળખાય છે, નિયુક્ત આલ્ફા, મ્યુ, Pi, Sigma, Theta, Omega, and Zeta.83�86 નોન-સ્ટ્રેસ્ડ પરિસ્થિતિઓ દરમિયાન, વર્ગ Mu અને Pi GST અનુક્રમે કિનાસિસ Ask1 અને JNK સાથે ક્રિયાપ્રતિક્રિયા કરે છે અને આ કિનાસિસને અટકાવે છે.87�89 એવું દર્શાવવામાં આવ્યું છે કે GSTP1 તેનાથી અલગ થઈ જાય છે ઓક્સિડેટીવ સ્ટ્રેસના પ્રતિભાવમાં JNK. 89 GSTP1 શારીરિક રીતે PRX VI સાથે પણ ક્રિયાપ્રતિક્રિયા કરે છે અને ઓક્સિડાઇઝ્ડ પ્રોટીનના ગ્લુટાથિઓનલેશન દ્વારા PRX એન્ઝાઇમ પ્રવૃત્તિની પુનઃપ્રાપ્તિ તરફ દોરી જાય છે. 90
Nonenzymatic એન્ટીઑકિસડન્ટોના
નોનેન્ઝાઇમેટિક એન્ટીઑકિસડન્ટોમાં ઓછા પરમાણુ-વજનના સંયોજનો, જેમ કે વિટામિન્સ (વિટામિન C અને E), બી-કેરોટિન, યુરિક એસિડ અને GSH, એક ટ્રિપેપ્ટાઇડ (Lg-glutamyl-L-cysteinyl-L-glycine)નો સમાવેશ થાય છે જેમાં થિયોલનો સમાવેશ થાય છે. સલ્ફાઇડ્રિલ) જૂથ.
વિટામિન સી (એસ્કર્બિક એસિડ)
પાણીમાં દ્રાવ્ય વિટામિન સી (એસ્કોર્બિક એસિડ) મુખ્યત્વે ઓક્સિજન મુક્ત રેડિકલને સાફ કરીને અંતઃકોશિક અને બાહ્યકોષીય જલીય-તબક્કાની એન્ટીઑકિસડન્ટ ક્ષમતા પ્રદાન કરે છે. તે વિટામીન E મુક્ત રેડિકલને વિટામીન E માં રૂપાંતરિત કરે છે. તેનું પ્લાઝ્મા સ્તર વય સાથે ઘટતું જોવા મળ્યું છે. 91,92
વિટામિન ઇ (એ-ટોકોફેરોલ)
લિપિડ-દ્રાવ્ય વિટામિન ઇ કોષ પટલની હાઇડ્રોફોબિક આંતરિક જગ્યામાં કેન્દ્રિત છે અને તે ઓક્સિડન્ટ-પ્રેરિત પટલની ઇજા સામે મુખ્ય સંરક્ષણ છે. વિટામિન ઇ પેરોક્સિલ રેડિકલને ઇલેક્ટ્રોનનું દાન કરે છે, જે લિપિડ પેરોક્સિડેશન દરમિયાન ઉત્પન્ન થાય છે. એ-ટોકોફેરોલ એ વિટામિન ઇનું સૌથી સક્રિય સ્વરૂપ છે અને કોષમાં મેમ્બ્રેન-બાઉન્ડ એન્ટીઑકિસડન્ટ છે. વિટામિન E કેન્સર કોશિકાઓના એપોપ્ટોસિસને ઉત્તેજિત કરે છે અને મુક્ત રેડિકલ રચનાઓને અટકાવે છે.93
ગ્લુટાથિઓન
જીએસએચ તમામ કોષ કમ્પાર્ટમેન્ટ્સમાં અત્યંત વિપુલ પ્રમાણમાં છે અને તે મુખ્ય દ્રાવ્ય એન્ટીઑકિસડન્ટ છે. જીએસએચ/જીએસએસજી રેશિયો ઓક્સિડેટીવ તણાવનું મુખ્ય નિર્ણાયક છે. GSH તેની એન્ટીઑકિસડન્ટ અસરોને ઘણી રીતે દર્શાવે છે. GSH તેના ઇલેક્ટ્રોનને H94O2 ને H2O અને O2 માં ઘટાડવા માટે દાન કરે છે. GSSG ને GSH રિડક્ટેઝ દ્વારા GSH માં ફરીથી ઘટાડી દેવામાં આવે છે જે ઇલેક્ટ્રોન દાતા તરીકે NAD(P)H નો ઉપયોગ કરે છે. GSH-Pxs લિપિડ પેરોક્સિડેશનથી કોષ પટલના રક્ષણ માટે પણ મહત્વપૂર્ણ છે. ઘટાડેલ ગ્લુટાથિઓન મેમ્બ્રેન લિપિડ્સમાં પ્રોટોનનું દાન કરે છે અને ઓક્સિડન્ટ હુમલાઓથી રક્ષણ આપે છે. 2
જીએસએચ એ જીએસએચ-પીએક્સ અને ટ્રાન્સફરઝ જેવા કેટલાક ડિટોક્સિફાઇંગ એન્ઝાઇમ માટે કોફેક્ટર છે. વિટામિન C અને E ને તેમના સક્રિય સ્વરૂપમાં ફેરવવામાં તેની ભૂમિકા છે. જીએસએચ પ્રોઆપોપ્ટોટિક અને એન્ટિએપોપ્ટોટિક સિગ્નલિંગ પાથવે સાથે ક્રિયાપ્રતિક્રિયા કરીને એપોપ્ટોસિસ સામે કોષોનું રક્ષણ કરે છે.
કેરોટીનોઈડ્સ (બી-કેરોટીન)
કેરોટીનોઈડ એ છોડમાં જોવા મળતા રંગદ્રવ્યો છે. પ્રાથમિક રીતે, બી-કેરોટીન પેરોક્સિલ (ROO), હાઇડ્રોક્સિલ (OH), અને સુપરઓક્સાઇડ (O22.) રેડિકલ સાથે પ્રતિક્રિયા કરે છે. સાંદ્રતા.96 કેરોટીનોઈડ્સ અને રેટિનોઈક એસિડ્સ (આરએ) બંને ટ્રાન્સક્રિપ્શન પરિબળોને નિયંત્રિત કરવામાં સક્ષમ છે. 97 બી-કેરોટીન ઓક્સિડન્ટ-પ્રેરિત NF-kB સક્રિયકરણ અને ઇન્ટરલ્યુકિન (IL)-98 અને ટ્યુમર નેક્રોસિસ પરિબળ-એ ઉત્પાદનને અટકાવે છે. કેરોટીનોઇડ્સ કોશિકાઓના એપોપ્ટોસિસને પણ અસર કરે છે. આરએની એન્ટિપ્રોલિફેરેટિવ અસરો કેટલાક અભ્યાસોમાં દર્શાવવામાં આવી છે. RA ની આ અસર મુખ્યત્વે રેટિનોઇક એસિડ રીસેપ્ટર્સ દ્વારા મધ્યસ્થી કરવામાં આવે છે અને કોષોના પ્રકારોમાં બદલાય છે. સ્તન્ય પ્રાણીઓમાં ગર્ભમાં રહેલા બચ્ચાની રક્ષા માટેનું આચ્છાદન કોશિકાઓમાં, રેટિનોઇક એસિડ રીસેપ્ટર સેલ ચક્ર ધરપકડ, એપોપ્ટોસિસ અથવા બંનેને પ્રેરિત કરીને વૃદ્ધિ અવરોધને ટ્રિગર કરવા માટે દર્શાવવામાં આવ્યું હતું.6
ઓક્સિડેટીવ સ્ટ્રેસની અસર: આનુવંશિક, ભૌતિકશાસ્ત્ર અને બાયોકેમિકલ મિકેનિઝમ્સ
ઓક્સિડેટીવ તણાવ ત્યારે થાય છે જ્યારે એન્ટીઑકિસડન્ટો અને ROS વચ્ચેનું સંતુલન ખોરવાય છે કારણ કે કાં તો એન્ટીઑકિસડન્ટોના ઘટાડાને કારણે અથવા ROS ના સંચયને કારણે. જ્યારે ઓક્સિડેટીવ તાણ થાય છે, ત્યારે કોષો ઓક્સિડન્ટ અસરોનો સામનો કરવાનો પ્રયાસ કરે છે અને રક્ષણાત્મક ઉત્સેચકો, ટ્રાન્સક્રિપ્શન પરિબળો અને માળખાકીય પ્રોટીનને એન્કોડિંગ જનીનોને સક્રિય કરીને અથવા મૌન કરીને રેડોક્સ સંતુલન પુનઃસ્થાપિત કરે છે. 101,102 ઓક્સિડાઇઝ્ડ અને ઘટાડેલા ગ્લુટાથિઓન (2GSH/GSSG) વચ્ચેનો ગુણોત્તર એક છે. શરીરમાં ઓક્સિડેટીવ તણાવના મહત્વના નિર્ણાયકોમાંથી. શરીરમાં આરઓએસનું ઊંચું ઉત્પાદન ડીએનએ માળખું બદલી શકે છે, પરિણામે પ્રોટીન અને લિપિડ્સમાં ફેરફાર થાય છે, ઘણા તણાવ-પ્રેરિત ટ્રાન્સક્રિપ્શન પરિબળો સક્રિય થાય છે, અને બળતરા તરફી અને બળતરા વિરોધી સાઇટોકીન્સનું ઉત્પાદન થાય છે.
ડીએનએ પર ઓક્સિડેટીવ તણાવની અસરો
આરઓએસ અનેક રીતે ડીએનએ ફેરફારો તરફ દોરી શકે છે, જેમાં પાયાના અધોગતિ, સિંગલ- અથવા ડબલ-સ્ટ્રેન્ડેડ ડીએનએ બ્રેક્સ, પ્યુરિન, પિરીમિડીન અથવા ખાંડ-બાઉન્ડ ફેરફારો, પરિવર્તન, કાઢી નાખવા અથવા ટ્રાન્સલોકેશન, અને પ્રોટીન સાથે ક્રોસ-લિંકિંગનો સમાવેશ થાય છે. આમાંના મોટાભાગના ડીએનએ ફેરફારો (ફિગ. 1) કાર્સિનોજેનેસિસ, વૃદ્ધત્વ અને ન્યુરોડિજનરેટિવ, કાર્ડિયોવેસ્ક્યુલર અને સ્વયંપ્રતિરક્ષા રોગો માટે અત્યંત સુસંગત છે. તમાકુનો ધુમાડો, રેડોક્સ ધાતુઓ અને નોનરેડોક્સ ધાતુઓ, જેમ કે આયર્ન, કેડમિયમ, ક્રોમ અને આર્સેનિક, મુક્ત રેડિકલ ઉત્પન્ન કરીને અથવા થિયોલ જૂથો સાથે બંધન કરીને કાર્સિનોજેનેસિસ અને વૃદ્ધત્વમાં પણ સામેલ છે. 8-OH-G ની રચના ઓક્સિડેટીવ તાણ દ્વારા થતી સૌથી જાણીતી ડીએનએ નુકસાન છે અને તે કાર્સિનોજેનેસિસ માટે સંભવિત બાયોમાર્કર છે.
જનીનોના પ્રમોટર પ્રદેશોમાં ટ્રાન્સક્રિપ્શન પરિબળો માટે સર્વસંમતિ ક્રમ હોય છે. આ ટ્રાન્સક્રિપ્શન ફેક્ટર-બાઇન્ડિંગ સાઇટ્સમાં જીસી-સમૃદ્ધ સિક્વન્સ હોય છે જે ઓક્સિડન્ટ હુમલાઓ માટે સંવેદનશીલ હોય છે. ટ્રાન્સક્રિપ્શન ફેક્ટર બંધનકર્તા સાઇટ્સમાં 8-OH-G DNA ની રચના ટ્રાન્સક્રિપ્શન પરિબળોના બંધનને સુધારી શકે છે અને આ રીતે સંબંધિત જનીનોની અભિવ્યક્તિને બદલી શકે છે જેમ કે AP-1 અને Sp-1 લક્ષ્ય ક્રમ માટે દર્શાવવામાં આવ્યું છે.103 8-OH-G ઉપરાંત, 8,59 -cyclo-29 -deoxyadenosine (cyclo-dA) પણ જો TATA બોક્સમાં સ્થિત હોય તો કોષ પ્રણાલીમાં રિપોર્ટર જનીનમાંથી ટ્રાન્સક્રિપ્શનને અવરોધે છે. . ટાટા-બંધનકર્તા પ્રોટીનનું બંધન સાયક્લો-ડીએની હાજરીથી ક્ષતિગ્રસ્ત થઈ શકે છે.
ઓક્સિડેટીવ તણાવ માઇક્રોસેટેલાઇટ (ટૂંકા ટેન્ડમ રિપીટ) પ્રદેશોની અસ્થિરતાનું કારણ બને છે. રેડોક્સ સક્રિય ધાતુના આયનો, હાઇડ્રોક્સિલ રેડિકલ માઇક્રોસેટેલાઇટની અસ્થિરતામાં વધારો કરે છે. 105 ઓક્સિડન્ટ ઇજાને કારણે એકલ-સ્ટ્રેન્ડેડ ડીએનએ વિરામ કોષો દ્વારા સરળતાથી સહન કરી શકાય તેમ હોવા છતાં, આયનાઇઝિંગ રેડિયેશન દ્વારા પ્રેરિત ડબલ-સ્ટ્રેન્ડેડ ડીએનએ વિરામ કોષના અસ્તિત્વ માટે નોંધપાત્ર જોખમ બની શકે છે.106
ડીએનએમાં સીપીજી ટાપુઓ પર મેથિલેશન એ એક મહત્વપૂર્ણ એપિજેનેટિક મિકેનિઝમ છે જેના પરિણામે જનીન શાંત થઈ શકે છે. 5-MeCyt થી 5-hydroxymethyl uracil (5-OHMeUra) નું ઓક્સિડેશન થાઇમિન અથવા 5-હાઈડ્રોક્સિમિથાઈલ સાયટોસિન મધ્યવર્તી ડિમિનેશન/ઓક્સિડેશન પ્રતિક્રિયાઓ દ્વારા થઈ શકે છે. ઓક્સિડેટીવ હુમલાઓ દ્વારા પ્રેરિત એબરન્ટ ડીએનએ મેથિલેશન પેટર્ન પણ ડીએનએ રિપેર પ્રવૃત્તિને અસર કરે છે.
લિપિડ્સ પર ઓક્સિડેટીવ તણાવની અસરો
ROS લિપિડ પેરોક્સિડેશનને પ્રેરિત કરી શકે છે અને મેમ્બ્રેન લિપિડ બાયલેયર ગોઠવણીને વિક્ષેપિત કરી શકે છે જે મેમ્બ્રેન-બાઉન્ડ રીસેપ્ટર્સ અને એન્ઝાઇમ્સને નિષ્ક્રિય કરી શકે છે અને પેશીઓની અભેદ્યતામાં વધારો કરી શકે છે. -લિંકેજ.109�110 112-હાઈડ્રોક્સી-4-નોનેનલ ઇન્ટ્રાસેલ્યુલર GSH ના ઘટાડાનું કારણ બને છે અને પેરોક્સાઇડ ઉત્પાદનને પ્રેરિત કરે છે,2 એપિડર્મલ ગ્રોથ ફેક્ટર રીસેપ્ટરને સક્રિય કરે છે,113,114 અને ફાઈબ્રોનેક્ટીન ઉત્પાદનને પ્રેરિત કરે છે. , ઓક્સિડેટીવ તાણના પરોક્ષ બાયોમાર્કર્સ તરીકે ઉપયોગમાં લેવાય છે, અને શ્વાસ બહાર કાઢવામાં આવતા શ્વસન કન્ડેન્સેટ અથવા બ્રોન્કોઆલ્વિઓલર લેવેજ પ્રવાહી અથવા ક્રોનિક અવરોધક પલ્મોનરી રોગના દર્દીઓ અથવા ધૂમ્રપાન કરનારાઓના ફેફસામાં વધેલા સ્તરો દર્શાવવામાં આવ્યા હતા.115�116
પ્રોટીન પર ઓક્સિડેટીવ તણાવની અસરો
આરઓએસ પેપ્ટાઈડ સાંકળના વિભાજન, પ્રોટીનના વિદ્યુત ચાર્જમાં ફેરફાર, પ્રોટીનનું ક્રોસ-લિંકિંગ અને ચોક્કસ એમિનો એસિડના ઓક્સિડેશનનું કારણ બની શકે છે અને તેથી ચોક્કસ પ્રોટીઝ દ્વારા અધોગતિ દ્વારા પ્રોટીઓલિસિસની સંવેદનશીલતામાં વધારો થાય છે. 120 પ્રોટીનમાં સિસ્ટીન અને મેથિઓનાઇન અવશેષો છે. ઓક્સિડેશન માટે ખાસ કરીને વધુ સંવેદનશીલ. 121 પ્રોટીનના સલ્ફહાઇડ્રેલ જૂથો અથવા મેથિઓનાઇન અવશેષોનું ઓક્સિડેશન રચનાત્મક ફેરફારો, પ્રોટીન પ્રગટ થવું અને અધોગતિનું કારણ બને છે. 8,121�123 ઉત્સેચકો કે જેમાં ધાતુઓ તેમની સક્રિય સાઇટ પર અથવા તેની નજીક હોય છે તે ખાસ કરીને ધાતુના ઉત્પ્રેરિત ઓક્સિડેશન પ્રત્યે વધુ સંવેદનશીલ હોય છે. ઉત્સેચકોના ઓક્સિડેટીવ ફેરફાર તેમની પ્રવૃત્તિઓને અવરોધે છે.124,125
કેટલાક કિસ્સાઓમાં, પ્રોટીનનું ચોક્કસ ઓક્સિડેશન થઈ શકે છે. ઉદાહરણ તરીકે, મેથિઓનાઇનને ઓક્સિડાઇઝ્ડ મેથિઓનાઇન સલ્ફોક્સાઇડ126 અને ફેનીલાલેનાઇનને ઓ-ટાયરોસિન 127 કરી શકાય છે; ડાયસલ્ફાઇડ બોન્ડ્સ બનાવવા માટે સલ્ફાઇડ્રિલ જૂથોને ઓક્સિડાઇઝ કરી શકાય છે; 128 અને કાર્બોનિલ જૂથો પ્રોટીનની બાજુની સાંકળોમાં દાખલ થઈ શકે છે. ગામા કિરણો, ધાતુ-ઉત્પ્રેરિત ઓક્સિડેશન, HOCl અને ઓઝોન કાર્બોનિલ જૂથોની રચનાનું કારણ બની શકે છે.129
સિગ્નલ ટ્રાન્સડક્શન પર ઓક્સિડેટીવ તણાવની અસરો
ROS સિગ્નલ ટ્રાન્સડક્શનમાં સામેલ કેટલાક જનીનોની અભિવ્યક્તિને પ્રેરિત કરી શકે છે. 1,130 GSH/GSSG માટે ઉચ્ચ ગુણોત્તર ઓક્સિડેટીવ નુકસાનથી કોષના રક્ષણ માટે મહત્વપૂર્ણ છે. આ ગુણોત્તરમાં વિક્ષેપ રેડોક્સ સંવેદનશીલ ટ્રાન્સક્રિપ્શન પરિબળોને સક્રિય કરે છે, જેમ કે એનએફ-કેબી, એપી-1, સક્રિય ટી કોશિકાઓના પરમાણુ પરિબળ અને હાયપોક્સિયા-ઇન્ડ્યુસિબલ પરિબળ 1, જે બળતરા પ્રતિભાવમાં સામેલ છે. ROS દ્વારા ટ્રાંસ્ક્રિપ્શન પરિબળોનું સક્રિયકરણ સિગ્નલ ટ્રાન્સડક્શન કાસ્કેડ્સ દ્વારા પ્રાપ્ત થાય છે જે માહિતીને બહારથી કોષની અંદર સુધી પ્રસારિત કરે છે. ટાયરોસિન કિનેઝ રીસેપ્ટર્સ, મોટાભાગના ગ્રોથ ફેક્ટર રીસેપ્ટર્સ, જેમ કે એપિડર્મલ ગ્રોથ ફેક્ટર રીસેપ્ટર, વેસ્ક્યુલર એન્ડોથેલિયલ ગ્રોથ ફેક્ટર રીસેપ્ટર અને પ્લેટલેટ ડેરિવ્ડ ગ્રોથ ફેક્ટર માટે રીસેપ્ટર, પ્રોટીન ટાયરોસિન ફોસ્ફેટેસીસ અને સેરીન/થ્રેઓનિન કિનાસીસ ROS131નું લક્ષ્ય છે. એક્સ્ટ્રા-સેલ્યુલર સિગ્નલ-રેગ્યુલેટેડ કિનાઝ, JNK, અને p133, જે મિટોજન-સક્રિય પ્રોટીન કિનાઝ પરિવારના સભ્યો છે અને પ્રસાર, ભિન્નતા અને એપોપ્ટોસિસ સહિત કોષમાં ઘણી પ્રક્રિયાઓમાં સામેલ છે, તે પણ ઓક્સિડન્ટ્સ દ્વારા નિયંત્રિત કરી શકાય છે.
ઓક્સિડેટીવ તણાવની સ્થિતિમાં, સી-જૂનના ડીએનએ-બંધન સ્થળમાં સિસ્ટીન અવશેષો, કેટલાક એપી-1 સબ્યુનિટ્સ અને અવરોધક kB કિનેઝ ઉલટાવી શકાય તેવા S-ગ્લુટાથિઓલેશનમાંથી પસાર થાય છે. ગ્લુટારેડોક્સિન અને TRX એ રેડોક્સ-સંવેદનશીલ સિગ્નલિંગ પાથવેઝના નિયમનમાં મહત્વની ભૂમિકા ભજવવાની જાણ કરવામાં આવી છે, જેમ કે NF-kB અને AP-1, p38 મિટોજન-સક્રિય પ્રોટીન કિનાઝ અને JNK.134�137
NF-kB ને ઓક્સિડેટીવ તણાવની સ્થિતિ, જેમ કે ROS, મુક્ત રેડિકલ અને યુવી ઇરેડિયેશનના પ્રતિભાવમાં સક્રિય કરી શકાય છે. IkB નું ફોસ્ફોરાયલેશન NF-kB ને મુક્ત કરે છે અને તેને જનીન ટ્રાન્સક્રિપ્શનને સક્રિય કરવા માટે ન્યુક્લિયસમાં પ્રવેશવાની મંજૂરી આપે છે. 138 સંખ્યાબંધ કિનાસ સેરીન અવશેષો પર ફોસ્ફોરીલેટ IkBs ને જાણ કરવામાં આવી હતી. આ કિનાસિસ NF-kB ના સક્રિયકરણ માટે ઓક્સિડેટીવ સંકેતોનું લક્ષ્ય છે. 139 ઘટાડતા એજન્ટો NF-kB DNA બંધનને વધારે છે, જ્યારે ઓક્સિડાઇઝિંગ એજન્ટો NF-kB ના DNA બંધનને અટકાવે છે. TRX NF-kB ના નિયમનમાં 140 વિરુદ્ધ ક્રિયાઓ કરી શકે છે: સાયટોપ્લાઝમમાં, તે IkB ના અધોગતિને અવરોધે છે અને NF-kB સક્રિયકરણને અટકાવે છે પરંતુ ન્યુક્લિયસમાં NF-kB DNA બંધનને વધારે છે. 2 ઓક્સિડેશન દ્વારા NF-kB નું સક્રિયકરણ- IkB ના ઘણા એન્ટીઑકિસડન્ટ સંરક્ષણ સંબંધિત જનીનો સક્રિયકરણમાં પરિણમે છે. NF-kB ઘણા જનીનોની અભિવ્યક્તિનું નિયમન કરે છે જે રોગપ્રતિકારક પ્રતિભાવમાં ભાગ લે છે, જેમ કે IL-141b, IL-1, ટ્યુમર નેક્રોસિસ ફેક્ટર-a, IL-6 અને કેટલાક સંલગ્ન પરમાણુઓ.8 NF-kB એન્જીયોજેનેસિસ અને પ્રસારને પણ નિયંત્રિત કરે છે અને કોષોનો તફાવત.
એપી-1 રેડોક્સ સ્ટેટ દ્વારા પણ નિયંત્રિત થાય છે. H2O2 ની હાજરીમાં, કેટલાક મેટલ આયનો AP-1 ના સક્રિયકરણને પ્રેરિત કરી શકે છે. GSH/GSSG ના ગુણોત્તરમાં વધારો એપી-1 બંધનકર્તાને વધારે છે જ્યારે GSSG એ AP-1.144 ના DNA બંધનને અટકાવે છે Fos/Jun heterodimer ના DNA બંધન દરેકના DNA-બંધનકર્તા ડોમેનમાં એક જ સંરક્ષિત સિસ્ટીનના ઘટાડાને કારણે વધે છે. પ્રોટીન,145 જ્યારે AP-1 નું DNA બંધન GSSG દ્વારા ઘણા સેલ પ્રકારોમાં અટકાવી શકાય છે, જે સૂચવે છે કે સિસ્ટીન અવશેષો દ્વારા ડિસલ્ફાઇડ બોન્ડની રચના AP-1 DNA બંધનને અટકાવે છે.
સમાપન
ઓક્સિડેટીવ તણાવ મેટાબોલિક પ્રતિક્રિયાઓ દ્વારા આરઓએસના વધુ ઉત્પાદનથી ઉદ્દભવી શકે છે જે ઓક્સિજનનો ઉપયોગ કરે છે અને વચ્ચેનું સંતુલન બદલી નાખે છે. ઓક્સિડન્ટ/એન્ટીઑકિસડન્ટ ઓક્સિડન્ટ્સની તરફેણમાં સ્થિતિ. ROS સેલ્યુલર મેટાબોલિક પ્રવૃત્તિઓ અને પર્યાવરણીય પરિબળો, જેમ કે હવા પ્રદૂષકો અથવા સિગારેટના ધુમાડા દ્વારા ઉત્પન્ન થાય છે. આરઓએસ એ અત્યંત પ્રતિક્રિયાશીલ પરમાણુઓ છે કારણ કે તેમની રચનામાં જોડી વગરના ઇલેક્ટ્રોન છે અને કોષમાં રહેલા કેટલાક જૈવિક મેક્રોમોલેક્યુલ્સ, જેમ કે કાર્બોહાઇડ્રેટ્સ, ન્યુક્લિક એસિડ્સ, લિપિડ્સ અને પ્રોટીન સાથે પ્રતિક્રિયા આપે છે અને તેમના કાર્યોમાં ફેરફાર કરે છે. આરઓએસ રેડોક્સ-સંવેદનશીલ ટ્રાન્સક્રિપ્શન પરિબળોના અપરેગ્યુલેશન દ્વારા અને હિસ્ટોન એસિટિલેશન/ડિસેટીલેશનમાં ફેરફાર દ્વારા ક્રોમેટિન રિમોડેલિંગ દ્વારા પણ કેટલાક જનીનોની અભિવ્યક્તિને અસર કરે છે. કોષની કાર્યક્ષમતા, સક્રિયકરણ, પ્રસાર અને અંગના કાર્ય માટે રેડોક્સ સ્થિતિનું નિયમન મહત્વપૂર્ણ છે.
સંદર્ભ
1. વાલ્કો એમ, રોડ્સ સીજે, મોનકોલ જે, ઇઝાકોવિક એમ, મઝુર એમ. ઓક્સિડેટીવ તણાવ-પ્રેરિત કેન્સરમાં મુક્ત રેડિકલ, ધાતુઓ અને એન્ટીઑકિસડન્ટો. કેમ બાયોલ ઇન્ટરેક્ટ. 2006;160:1�40.
2. હેલીવેલ બી, ગુટેરિજ જેએમસી. જીવવિજ્ઞાન અને દવામાં મુક્ત રેડિકલ. 3જી આવૃત્તિ. ન્યૂ યોર્ક: ઓક્સફોર્ડ યુનિવર્સિટી પ્રેસ; 1999.
3. માર્નેટ એલજે. લિપિડ પેરોક્સિડેશન ડીએનએ મેલોન્ડિઆલ્ડિહાઇડ દ્વારા નુકસાન. Mutat Res. 1999;424:83�95.
4. Siems WG, Grune T, Esterbauer H. 4-Hydroxynonenal formation દરમિયાન ઇસ્કેમિયા અને ઉંદરના નાના આંતરડાના રિપરફ્યુઝન. જીવન વિજ્ઞાન. 1995;57:785�789.
5. સ્ટેડટમેન ER. વૃદ્ધત્વમાં ઓક્સિડન્ટ પ્રજાતિઓની ભૂમિકા. કર મેડ કેમ. 2004;11:1105�1112.
6. વાંગ MY, ઢીંગરા કે, હિટ્ટેલમેન ડબલ્યુએન, લિહર જેજી, ડીએન્ડ્રેડ એમ, લી ડીએચ. લિપિડ પેરોક્સિડેશન-પ્રેરિત પુટેટિવ મેલોન્ડિઆલ્ડિહાઇડ-ડીએનએ માનવ સ્તનના પેશીઓમાં એડક્ટ કરે છે. કેન્સર એપિડેમિઓલ બાયોમાર્કર્સ અગાઉના. 1996;5:705�710.
7. જેનર પી. પાર્કિન્સન રોગમાં ઓક્સિડેટીવ તણાવ. એન ન્યુરોલ. 2003;53: S26�S36.
8. લિરાસ એલ, કેર્ન્સ એનજે, જેનર એ, જેનર પી, હેલીવેલ બી. અલ્ઝાઈમર રોગ ધરાવતા દર્દીઓના મગજમાં પ્રોટીન, લિપિડ્સ અને ડીએનએને ઓક્સિડેટીવ નુકસાનનું મૂલ્યાંકન. જે ન્યુરોકેમ. 1997;68:2061�2069.
9. સેરે એલએમ, સ્મિથ એમએ, પેરી જી. રસાયણશાસ્ત્ર અને ન્યુરોડિજનરેટિવ રોગમાં ઓક્સિડેટીવ સ્ટ્રેસની બાયોકેમિસ્ટ્રી. કર મેડ કેમ. 2001;8:721�738.
10. તોશનીવાલ પીકે, ઝરલિંગ ઇજે. મલ્ટિપલ સ્ક્લેરોસિસમાં લિપિડ પેરોક્સિડેશનમાં વધારો થવાના પુરાવા. ન્યુરોકેમ રેસ. 1992;17:205�207.
11. ધલ્લા એનએસ, ટેમસાહ આરએમ, નેટિકાડન ટી. કાર્ડિયોવેસ્ક્યુલર રોગોમાં ઓક્સિડેટીવ સ્ટ્રેસની ભૂમિકા. જે હાઇપરટેન્સ. 2000;18:655�673.
12. કાસ્પારોવા એસ, બ્રેઝોવા વી, વાલ્કો એમ, હોરેકી જે, મલિનરિક વી, એટ અલ. ક્રોનિક મગજ હાયપોપરફ્યુઝનના ઉંદર મોડેલમાં ઓક્સિડેટીવ તણાવનો અભ્યાસ. ન્યુરોકેમ ઇન્ટ. 2005;46:601�611.
13. કેર એસ, બ્રોસ્નન એમજે, મેકઇન્ટાયર એમ, રીડ જેએલ, ડોમિનિકઝાક એએફ, હેમિલ્ટન સીએ. આનુવંશિક હાયપરટેન્શનના મોડેલમાં સુપરઓક્સાઇડ આયનોનું ઉત્પાદન વધે છે: એન્ડોથેલિયમની ભૂમિકા. હાયપરટેન્શન. 1999;33:1353�1358.
14. કુકરેજા આરસી, હેસ એમએલ. ઓક્સિજન ફ્રી-રેડિકલ સિસ્ટમ: મેમ્બ્રેન-પ્રોટીન ક્રિયાપ્રતિક્રિયાઓ દ્વારા સમીકરણોથી કાર્ડિયોવેસ્ક્યુલર ઇજા અને રક્ષણ સુધી. કાર્ડિયોવેસ્ક રેસ. 1992;26:641�655.
15. Asami S, Manabe H, Miyake J, Tsurudome Y, Hirano T, et al. સિગારેટનું ધૂમ્રપાન માનવ ફેફસાની મધ્યસ્થ જગ્યામાં ઓક્સિડેટીવ ડીએનએ નુકસાન, 8-હાઈડ્રોક્સાઈડિયોક્સીગુઆનોસિન,માં વધારો કરે છે. કાર્સિનોજેનેસિસ. 1997;18:1763�1766.
16. એન્ડ્રીડિસ એએ, હેઝન એસએલ, કોમહેર એસએ, એર્ઝુરમ એસસી. અસ્થમામાં ઓક્સિડેટીવ અને નાઈટ્રોસેટીવ ઘટનાઓ. ફ્રી રેડિક બાયોલ મેડ. 2003;35:213�225.
17. Comhair SA, Ricci KS, Arroliga M, Lara AR, Dweik RA, et al. અસ્થમામાં એરફ્લો અવરોધ સાથે પ્રણાલીગત સુપરઓક્સાઇડ ડિસમ્યુટેઝની ઉણપનો સહસંબંધ. એમ જે રેસ્પિર ક્રિટ કેર મેડ. 2005;172:306�313.
18. Comhair SA, Xu W, Ghosh S, Thunnissen FB, Almasan A, et al. અસ્થમાના એરવે રિમોડેલિંગ અને પ્રતિક્રિયાશીલતાના પેથોફિઝિયોલોજીમાં સુપરઓક્સાઇડ ડિસમ્યુટેઝ નિષ્ક્રિયતા. એમ જે પથોલ. 2005;166:663�674.
19. Dut R, Dizdar EA, Birben E, Sackesen C, Soyer OU, Besler T, Kalayci O. ઓક્સિડેટીવ તણાવ અને અસ્થમાવાળા બાળકોના વાયુમાર્ગમાં તેના નિર્ધારકો. એલર્જી. 2008;63:1605�1609.
20. Ercan H, Birben E, Dizdar EA, Keskin O, Karaaslan C, et al. ઓક્સિડેટીવ તાણ અને બાળપણના અસ્થમામાં ઓક્સિડન્ટ ઇજાના આનુવંશિક અને રોગચાળાના નિર્ધારકો. જે એલર્જી ક્લિન ઇમ્યુનોલ. 2006;118:1097�1104.
21. Fitzpatrick AM, Teague WG, Holguin F, Yeh M, Brown LA. ગંભીર અસ્થમા સંશોધન કાર્યક્રમ. ગંભીર અસ્થમાવાળા બાળકોમાં એરવે ગ્લુટાથિઓન હોમિયોસ્ટેસિસ બદલાય છે: ઓક્સિડન્ટ તણાવ માટેના પુરાવા. જે એલર્જી ક્લિન ઇમ્યુનોલ. 2009;123:146�152.
22. મિલર ડીએમ, બ્યુટનર જીઆર, ઓસ્ટ એસડી. "ઓટોક્સિડેશન" પ્રતિક્રિયાઓના ઉત્પ્રેરક તરીકે સંક્રમણ ધાતુઓ. ફ્રી રેડિક બાયોલ મેડ. 1990;8:95�108.
23. Dupuy C, Virion A, Ohayon R, Kaniewski J, D�me D, Pommier J. થાઇરોઇડ પ્લાઝ્મા મેમ્બ્રેનમાં NADPH ઓક્સિડેઝ દ્વારા ઉત્પ્રેરિત હાઇડ્રોજન પેરોક્સાઇડ રચનાની પદ્ધતિ. જે બાયોલ કેમ. 1991;266:3739�3743.
24. ગ્રેન્જર ડીએન. ઇસ્કેમિયારેપરફ્યુઝન ઇજામાં ઝેન્થાઇન ઓક્સિડેઝ અને ગ્રાન્યુલોસાઇટ્સની ભૂમિકા. એમ જે ફિઝિયોલ. 1988;255:H1269�H1275.
25. ફેન્ટન HJH. આયર્નની હાજરીમાં ટાર્ટરિક એસિડનું ઓક્સિડેશન. જે કેમ સોસી. 1984;65:899�910.
26. હેબર એફ, વેઇસ જેજે. આયર્ન ક્ષાર દ્વારા હાઇડ્રોજન પેરોક્સાઇડનું ઉત્પ્રેરક વિઘટન. પ્રોક આર સોક લંડ સેર એ. 1934;147:332�351.
27. લિઓચેવ એસઆઈ, ફ્રિડોવિચ I. હેબર વેઈસ 70 વર્ષ પછી સાયકલ ચલાવે છે: એક વૈકલ્પિક દૃશ્ય. રેડોક્સ રેપ. 2002;7:55�57.
28. Klebanoff SJ. માયલોપેરોક્સિડેઝ: મિત્ર અને શત્રુ. જે લ્યુકોક બાયોલ. 2005;77:598�625.
29. વ્હાઇટમેન એમ, જેનર એ, હેલીવેલ બી. આઇસોલેટેડ કાફ થાઇમસ ડીએનએમાં હાઇપોક્લોરસ એસિડ-પ્રેરિત આધાર ફેરફારો. કેમ રેસ ટોક્સિકોલ. 1997;10:1240�1246.
30. કુલચરીક પીએ, હેનેકે જેડબ્લ્યુ. માનવ ફેગોસાઇટ્સની માયલોપેરોક્સિડેઝ સિસ્ટમ દ્વારા ઉત્પાદિત હાઇપોક્લોરસ એસિડ ડીએનએ અને પ્રોટીન વચ્ચે સહસંયોજક ક્રોસ-લિંકને પ્રેરિત કરે છે. બાયોકેમિસ્ટ્રી. 2001;40:3648�3656.
31. બ્રેનન ML, Wu W, Fu X, Shen Z, Song W, et al. બે વિવાદોની વાર્તા: વિવોમાં ઇઓસિનોફિલ પેરોક્સિડેઝ અને માયલોપેરોક્સિડેસની ખામીયુક્ત ઉંદરનો ઉપયોગ કરીને નાઇટ્રોટાયરોસિન રચનામાં પેરોક્સિડેઝની ભૂમિકા અને પેરોક્સિડેઝ-જનરેટ રિએક્ટિવ નાઇટ્રોજન પ્રજાતિઓની પ્રકૃતિ બંનેને વ્યાખ્યાયિત કરવી. જે બાયોલ કેમ. 2002;277:17415�17427.
32. ડેન્ઝલર કેએલ, બોર્ચર્સ એમટી, ક્રોસબી જેઆર, સિસ્લેવિઝ જી, હાઈન્સ ઈએમ, એટ અલ. વ્યાપક ઇઓસિનોફિલ ડિગ્રેન્યુલેશન અને એરવે પ્રોટીનનું પેરોક્સિડેઝ-મધ્યસ્થ ઓક્સિડેશન પલ્મોનરી ઇન્ફ્લેમેશનના માઉસ ઓવલબ્યુમિન-ચેલેન્જ મોડેલમાં થતું નથી. જે ઇમ્યુનોલ. 2001;167:1672�1682.
33. વાન ડેલેન સીજે, વિન્ટરબોર્ન સીસી, સેંથિલમોહન આર, કેટલ એજે. નાઇટ્રાઇટ સબસ્ટ્રેટ અને માયલોપેરોક્સિડેઝના અવરોધક તરીકે. બળતરાના સ્થળો પર નાઈટ્રેશન અને હાયપોક્લોરસ એસિડ ઉત્પાદન માટે અસરો. જે બાયોલ કેમ. 2000;275:11638�11644.
34. વુડ એલજી, ફિટ્ઝગેરાલ્ડ ડીએ, ગિબ્સન પીજી, કૂપર ડીએમ, ગર્ગ એમએલ. પ્લાઝ્મા આઇસોપ્રોસ્ટેનેસ દ્વારા નિર્ધારિત લિપિડ પેરોક્સિડેશન હળવા અસ્થમામાં રોગની તીવ્રતા સાથે સંબંધિત છે. લિપિડ્સ. 2000;35:967�974.
35. મોન્ટુચી પી, કોરાડી એમ, સિયાબેટોની જી, નાઈટીંગેલ જે, ખારીટોનોવ એસએ, બાર્નેસ પીજે. 8-આઇસોપ્રોસ્ટેન વધારો, જે ઓક્સિડેટીવ તણાવનું માર્કર છે, અસ્થમાના દર્દીઓના શ્વાસ બહાર કાઢતા કન્ડેન્સેટમાં. એમ જે રેસ્પિર ક્રિટ કેર મેડ. 1999;160:216�220.
36. ચર્ચ DF, Pryor WA. સિગારેટના ધુમાડાની મુક્ત-આમૂલ રસાયણશાસ્ત્ર અને તેની ઝેરી અસરો. પર્યાવરણ આરોગ્ય પરિપ્રેક્ષ્ય. 1985;64:111�126.
37. હિલ્ટરમેન જેટી, લેપરે ટીએસ, વેન બ્રી એલ, સ્ટીરેનબર્ગ પીએ, બ્રાહિમ જેજે, એટ અલ. અસ્થમાના રોગમાંથી ગળફામાં અને શ્વાસનળીના લેવેજ પ્રવાહીમાં ઓઝોન-પ્રેરિત બળતરાનું મૂલ્યાંકન: વાયુ પ્રદૂષણ અને અસ્થમા પર રોગચાળાના અભ્યાસમાં એક નવું બિન-આક્રમક સાધન. ફ્રી રેડિક બાયોલ મેડ. 1999;27:1448�1454.
38. નાઇટીંગેલ જેએ, રોજર્સ ડીએફ, બાર્નેસ પીજે. સામાન્ય અને અસ્થમાના વિષયોમાં શ્વાસમાં લેવાયેલા નાઈટ્રિક ઑક્સાઈડ, પલ્મોનરી ફંક્શન અને પ્રેરિત ગળફા પર શ્વાસમાં લેવાયેલા ઓઝોનની અસર. થોરાક્સ. 1999;54:1061�1069.
39. Cho AK, Sioutas C, Miguel AH, Kumagai Y, Schmitz DA, et al. લોસ એન્જલસ બેસિનમાં વિવિધ સ્થળોએ એરબોર્ન પાર્ટિક્યુલેટ મેટરની રેડોક્સ પ્રવૃત્તિ. પર્યાવરણ Res. 2005;99:40�47.
40. કોમહેર એસએ, થોમસેન એમજે, એર્ઝુરમ એસસી. 2% O(100) અથવા સિગારેટના ધુમાડાના સંપર્કમાં આવતા સ્વસ્થ વ્યક્તિઓના વાયુમાર્ગમાં એક્સ્ટ્રા સેલ્યુલર ગ્લુટાથિઓન પેરોક્સિડેઝ અને નાઈટ્રિક ઓક્સાઇડ સિન્થેઝ 2નું વિભેદક ઇન્ડક્શન. એમ જે રેસ્પિર સેલ મોલ બાયોલ. 2000;23:350�354.
41. મૈથે એમએ, ગીઝર ટી, મેટાલોન એસ, ઇશિરોપોલોસ એચ. તીવ્ર શ્વસન તકલીફ સિન્ડ્રોમમાં ઓક્સિડન્ટ-મધ્યસ્થી ફેફસાની ઇજા. ક્રિટ કેર મેડ. 1999;27:2028�2030.
42. બાયગ્લો જેઇ, મિશેલ જેબી, હેલ્ડ કે. એક્સ-રે પ્રતિભાવમાં પેરોક્સાઇડ અને સુપરઓક્સાઇડનું મહત્વ. ઇન્ટ જે રેડિયેટ ઓન્કોલ બાયોલ ફિઝ. 1992;22:665�669.
43. Chiu SM, Xue LY, Friedman LR, Oleinick NL. આયનાઇઝિંગ રેડિયેશન માટે ન્યુક્લિયર મેટ્રિક્સ એટેચમેન્ટ સાઇટ્સનું કોપર આયન-મધ્યસ્થી સંવેદનશીલતા. બાયોકેમિસ્ટ્રી. 1993;32:6214�6219.
44. નારાયણન પીકે, ગુડવિન EH, લેહનર્ટ BE. આલ્ફા કણો માનવ કોષોમાં સુપરઓક્સાઇડ આયન અને હાઇડ્રોજન પેરોક્સાઇડનું જૈવિક ઉત્પાદન શરૂ કરે છે. કેન્સર રે. 1997;57:3963�3971.
45. ટટલ SW, Varnes ME, Mitchell JB, Biaglow JE. ઓક્સિડેટીવ પેન્ટોઝ ચક્ર પ્રવૃત્તિમાં ઉણપ ધરાવતા CHO સેલ લાઇન્સમાં રાસાયણિક ઓક્સિડન્ટ્સ અને રેડિયેશન પ્રત્યે સંવેદનશીલતા. ઇન્ટ જે રેડિયેટ ઓન્કોલ બાયોલ ફિઝ. 1992;22: 671�675.
46. ગુઓ જી, યાન-સેન્ડર્સ વાય, લિન-કુક બીડી, વાંગ ટી, ટામે ડી, એટ અલ. મેંગેનીઝ
કિરણોત્સર્ગ પ્રેરિતમાં સુપરઓક્સાઇડ ડિસમ્યુટેઝ-મધ્યસ્થિત જનીન અભિવ્યક્તિ
અનુકૂલનશીલ પ્રતિભાવો. મોલ સેલ બાયોલ. 2003;23:2362�2378.
47. Azzam EI, de Toledo SM, Spitz DR, Little JB. ઓક્સિડેટીવ ચયાપચય
બાયસ્ટેન્ડરમાં સિગ્નલ ટ્રાન્સડક્શન અને માઇક્રોન્યુક્લિયસ રચનાને મોડ્યુલેટ કરે છે
એ-પાર્ટીકલમાંથી કોષો સામાન્ય માનવ ફાઇબ્રોબ્લાસ્ટને ઇરેડિયેટ કરે છે. કેન્સર Res.
2002;62:5436�5442.
48. લીચ જેકે, વેન તુયલ જી, લિન પીએસ, શ્મિટ-ઉલ્રિચ આર, મિકેલસન આરબી.
આયનાઇઝિંગ રેડિયેશન-પ્રેરિત, મિટોકોન્ડ્રિયા-આશ્રિત પ્રતિક્રિયાશીલ પેઢી
ઓક્સિજન/નાઇટ્રોજન. કેન્સર રે. 2001;61:3894�3901.
49. ડેન્ટ પી, યાકૂબ એ, ફિશર પીબી, હેગન એમપી, ગ્રાન્ટ એસ. એમએપીકે પાથવેઝ ઇન
રેડિયેશન પ્રતિક્રિયાઓ. ઓન્કોજીન. 2003;22:5885�5896.
50. વેઇ એસજે, બોટેરો એ, હિરોટા કે, બ્રેડબરી સીએમ, માર્કોવિના એસ, એટ અલ. થિયોરેડોક્સિન
પરમાણુ સ્થાનાંતરણ અને રેડોક્સ પરિબળ-1 સાથેની ક્રિયાપ્રતિક્રિયા આયનાઇઝિંગ રેડિયેશનના પ્રતિભાવમાં AP-1 ટ્રાન્સક્રિપ્શન પરિબળને સક્રિય કરે છે. કેન્સર રે. 2000;60:6688�6695.
51. કેડેટ J, Douki T, Gasparutto D, Ravanat JL. ડીએનએને ઓક્સિડેટીવ નુકસાન: રચના, માપન અને બાયોકેમિકલ લક્ષણો. Mutat Res. 2003;531:5�23.
52. યોકોયા એ, કનિફ એસએમ, ઓ�નીલ પી. ગેમરેડિયેશન દ્વારા પ્લાઝમિડ ડીએનએ ફિલ્મોમાં સ્ટ્રાન્ડ બ્રેક્સ અને બેઝ લેઝનના ઇન્ડક્શન પર હાઇડ્રેશનની અસર. J Am Chem Soc. 2002;124:8859�8866.
53. Janssen YM, વેન Houten B, Borm PJ, Mossman BT. ઓક્સિડેટીવ નુકસાન માટે કોષ અને પેશીઓની પ્રતિક્રિયાઓ. લેબ રોકાણ. 1993;69:261�274.
54. ઇવાનાગા એમ, મોરી કે, આઇડા ટી, ઉરાતા વાય, માત્સુઓ ટી, એટ અલ. T98G માનવ ગ્લિઓબ્લાસ્ટોમા કોષોમાં આયનાઇઝિંગ રેડિયેશન દ્વારા ગામા ગ્લુટામિલસિસ્ટીન સિન્થેટેઝનું ન્યુક્લિયર ફેક્ટર કપ્પા બી આધારિત ઇન્ડક્શન. ફ્રી રેડિક બાયોલ મેડ. 1998;24:1256�1268.
55. Stohs SJ, Bagchi D. ધાતુના આયનોની ઝેરીતામાં ઓક્સિડેટીવ મિકેનિઝમ્સ. ફ્રી રેડિક બાયોલ મેડ. 1995;18:321�336.
56. લિયોનાર્ડ એસએસ, હેરિસ જીકે, શી એક્સ. મેટલ-પ્રેરિત ઓક્સિડેટીવ સ્ટ્રેસ અને સિગ્નલ ટ્રાન્સડક્શન. ફ્રી રેડિક બાયોલ મેડ. 2004;37:1921�1942.
57. શી એચ, શી એક્સ, લિયુ કેજે. આર્સેનિક ટોક્સિસિટી અને કાર્સિનોજેનેસિસની ઓક્સિડેટીવ મિકેનિઝમ. મોલ સેલ બાયોકેમ. 2004;255:67�78.
58. Pi J, Horiguchi S, Sun Y, Nikaido M, Shimojo N, Hayashi T. સસલામાં આર્સેનેટના લાંબા સમય સુધી મૌખિક સંપર્કને કારણે નાઈટ્રિક ઑકસાઈડની રચનામાં ક્ષતિ માટે સંભવિત પદ્ધતિ. ફ્રી રેડિક બાયોલ મેડ.2003;35:102�113.
59. Rin K, Kawaguchi K, Yamanaka K, Tezuka M, Oku N, Okada S. DNAstrand બ્રેક્સ ડાયમેથાઈલરસિનિક એસિડ દ્વારા પ્રેરિત, અકાર્બનિક આર્સેનિકના મેટાબોલાઇટ, સુપરઓક્સાઈડ એનિઓન રેડિકલ દ્વારા મજબૂત રીતે વધારે છે. બાયોલ ફાર્મ બુલ. 1995;18:45�58.
60. વાલ્કેસ એમપી, લિયુ જે, વોર્ડ જેએમ, દિવાન એલએ. આર્સેનિક કાર્સિનોજેનેસિસ અંતર્ગત મિકેનિઝમ્સ: સગર્ભાવસ્થા દરમિયાન અકાર્બનિક આર્સેનિકના સંપર્કમાં આવતા ઉંદરની અતિસંવેદનશીલતા. ટોક્સિકોલોજી. 2004;198:31�38.
61. શિલર સીએમ, ફાઉલર બીએ, વુડ્સ જેએસ. પિરુવેટ ડિહાઇડ્રોજેનેઝ સક્રિયકરણ પર આર્સેનિકની અસરો. પર્યાવરણ આરોગ્ય પરિપ્રેક્ષ્ય. 1977;19:205�207.
62. મોન્ટેરિયો એચપી, બેચારા ઇજેએચ, અબ્દલ્લા ડીએસપી. ન્યુરોલોજીકલ પોર્ફિરિયા અને લીડ પોઈઝનીંગમાં ફ્રી રેડિકલની સંડોવણી. મોલ સેલ બાયોકેમ. 1991;103:73�83.
63. ત્રિપાઠી આર.એમ., રઘુનાથ આર, મહાપાત્રા એસ. બ્લડ લીડ અને બાળકોના Cd, Cu, Zn, Fe અને હિમોગ્લોબિન સ્તરો પર તેની અસર. વિજ્ઞાન કુલ પર્યાવરણ. 2001;277:161�168.
64. નેહરુ બી, દુઆ આર. લીડ ન્યુરોટોક્સિસિટી પર ડાયેટરી સેલેનિયમની અસર. જે એન્વાયરોન પેથોલ ટોક્સિકોલ ઓન્કોલ. 1997; 16:47�50.
65. રીડ TM, Feig DI, Loeb LA. મેટલ-પ્રેરિત ઓક્સિજન રેડિકલ દ્વારા મ્યુટાજેનેસિસ. પર્યાવરણ આરોગ્ય પરિપ્રેક્ષ્ય. 1994;102(પુરવઠા 3):57�61.
66. કિનુલા વીએલ, ક્રેપો જેડી. ફેફસાં અને માનવ ફેફસાના રોગોમાં સુપરઓક્સાઇડ છૂટી જાય છે. એમ જે રેસ્પિર ક્રિટ કેર મેડ. 2003;167:1600�1619.
67. કિનુલા વીએલ. માનવ ફેફસામાં બળતરા અવસ્થાઓ દરમિયાન ઓક્સિજન મેટાબોલાઇટ્સનું ઉત્પાદન અને અધોગતિ. કર ડ્રગ ઇન્ફ્લેમ એલર્જીને ટાર્ગેટ કરે છે. 2005;4:465�470.
68. Zelko IN, Mariani TJ, Folz RJ. સુપરઓક્સાઇડ ડિસમ્યુટેઝ મલ્ટિજીન ફેમિલી: CuZn-SOD (SOD1), Mn-SOD (SOD2), અને EC-SOD (SOD3) જનીન રચનાઓ, ઉત્ક્રાંતિ અને અભિવ્યક્તિની સરખામણી. ફ્રી રેડિક બાયોલ મેડ. 2002;33:337�349.
69. કિર્કમેન એચએન, રોલ્ફો એમ, ફેરારિસ એએમ, ગેતાની જીએફ. NADPH દ્વારા કેટાલેઝના રક્ષણની પદ્ધતિઓ. ગતિશાસ્ત્ર અને સ્ટોઇકોમેટ્રી. જે બાયોલ કેમ. 1999;274:13908�13914.
70. Floh� L. Glutathione peroxidase. મૂળભૂત જીવન વિજ્ઞાન. 1988;49:663�668.
71. આર્થર જેઆર. ગ્લુટાથિઓન પેરોક્સિડેઝ. સેલ મોલ લાઇફ સાય. 2000;57:1825�1835.
72. Chu FF, Doroshow JH, Esworthy RS. નવા સેલ્યુલર સેલેનિયમ-આશ્રિત ગ્લુટાથિઓન પેરોક્સિડેઝ, GSHPx-GI ની અભિવ્યક્તિ, લાક્ષણિકતા અને પેશીઓનું વિતરણ. જે બાયોલ કેમ. 1993;268:2571�2576.
73. Comhair SA, Bhathena PR, Farver C, Thunnissen FB, Erzurum SC. અસ્થમાના ફેફસામાં એક્સ્ટ્રા સેલ્યુલર ગ્લુટાથિઓન પેરોક્સિડેઝ ઇન્ડક્શન: માનવ વાયુમાર્ગ ઉપકલા કોષોમાં અભિવ્યક્તિના રેડોક્સ નિયમન માટેના પુરાવા. FASEB જે. 2001; 15:70�78.
74. ગ્રોમર એસ, ઉરીગ એસ, બેકર કે. સાયન્સથી ક્લિનિક સુધી થિયોરેડોક્સિન સિસ્ટમ. મેડ રેવ. 2004;24:40�89.
75. કિન્નુલા વીએલ, લેહટોનન એસ, કાર્ટીનાહો-વિક આર, લકરી ઇ, પી��ક્ક� પી, એટ અલ. માનવ ફેફસાં અને પલ્મોનરી સાર્કોઇડોસિસમાં પેરોક્સિરેડોક્સિનની કોષ વિશિષ્ટ અભિવ્યક્તિ. થોરાક્સ. 2002;57:157�164.
76. ડુબ્યુસન એમ, વેન્ડર સ્ટ્રિક્ટ ડી, ક્લિપ એ, એટીન એફ, નૌસર ટી, એટ અલ. હ્યુમન પેરોક્સિરેડોક્સિન 5 એ પેરોક્સિનાઈટ્રેટ રીડક્ટેઝ છે. FEBS લેટ. 2004;571:161�165.
77. હોલ્મગ્રેન A. થિયોરેડોક્સિન અને ગ્લુટેરેડોક્સિન સિસ્ટમ્સનું એન્ટીઑકિસડન્ટ કાર્ય. એન્ટિઓક્સિડ રેડોક્સ સિગ્નલ. 2000;2:811�820.
78. ડિકિન્સન ડીએ, ફોરમેન એચજે. સંરક્ષણ અને સિગ્નલિંગમાં ગ્લુટાથિઓન: નાના થિયોલમાંથી પાઠ. એન NY Acad Sci. 2002;973:488�504.
79. Sies H. Glutathione અને સેલ્યુલર કાર્યોમાં તેની ભૂમિકા. ફ્રી રેડિક બાયોલ મેડ. 1999;27:916�921.
80. લેડનર જેઇ, પાર્સન્સ જેએફ, રાઇફ સીએલ, ગિલીલેન્ડ જીએલ, આર્મસ્ટ્રોંગ આરએન. ગ્લુટાથિઓન ટ્રાન્સફરસેસ માટે સમાંતર ઉત્ક્રાંતિ માર્ગો: મિટોકોન્ડ્રીયલ ક્લાસ કપ્પા એન્ઝાઇમ rGSTK1-1નું માળખું અને પદ્ધતિ. બાયોકેમિસ્ટ્રી. 2004;43:52�61.
81. રોબિન્સન એ, હટલી જીએ, બૂથ એચએસ, બોર્ડ પીજી. હ્યુમન કપ્પા ક્લાસ ગ્લુટાથિઓન ટ્રાન્સફરસેસના મોડેલિંગ અને બાયોઇન્ફોર્મેટિક્સ અભ્યાસો પ્રોકેરીયોટિક 2-હાઈડ્રોક્સીક્રોમીન-2-કાર્બોક્સિલેટ આઇસોમેરેસીસ માટે હોમોલોજી સાથે નવલકથા ત્રીજા ટ્રાન્સફરસેસ પરિવારની આગાહી કરે છે. બાયોકેમ જે. 2004;379:541�552.
82. જેકોબસન પીજે, મોર્ગેનસ્ટર્ન આર, મેનસિની જે, ફોર્ડ-હચિન્સન એ, પર્સન બી. એમએપીઇજીડાની સામાન્ય માળખાકીય વિશેષતાઓ ઇકોસાનોઇડ અને ગ્લુટાથિઓન ચયાપચયમાં અત્યંત વૈવિધ્યસભર કાર્યો સાથે મેમ્બ્રેન સંબંધિત પ્રોટીનની વ્યાપક સુપરફેમિલી. પ્રોટીન સાય. 1999;8:689�692.
83. Hayes JD, Pulford DJ. ગ્લુટાથિઓન એસ-ટ્રાન્સફેરેઝ સુપરજીન ફેમિલી: જીએસટીનું નિયમન અને કેન્સર કેમોપ્રોટેક્શન અને ડ્રગ રેઝિસ્ટન્સમાં આઇસોએન્ઝાઇમ્સનું યોગદાન. ક્રિટ રેવ બાયોકેમ મોલ બાયોલ. 1995;30:445�600.
84. આર્મસ્ટ્રોંગ આરએન. ગ્લુટાથિઓન ટ્રાન્સફરસેસનું માળખું, ઉત્પ્રેરક પદ્ધતિ અને ઉત્ક્રાંતિ. કેમ રેસ ટોક્સિકોલ. 1997;10:2�18.
85. હેયસ જેડી, મેકલેલન એલઆઈ. ગ્લુટાથિઓન અને ગ્લુટાથિઓન-આશ્રિત ઉત્સેચકો ઓક્સિડેટીવ તણાવ સામે સંકલિત રીતે નિયંત્રિત સંરક્ષણનું પ્રતિનિધિત્વ કરે છે. ફ્રી રેડિક રેસ. 1999;31:273�300.
86. શીહાન ડી, મીડે જી, ફોલી વીએમ, ડાઉડ સીએ. ગ્લુટાથિઓન ટ્રાન્સફરસેસનું માળખું, કાર્ય અને ઉત્ક્રાંતિ: એક પ્રાચીન એન્ઝાઇમ સુપરફેમિલીના બિનસસ્તન સભ્યોના વર્ગીકરણ માટે સૂચિતાર્થ. બાયોકેમ જે. 2001;360:1�16.
87. Cho SG, Lee YH, Park HS, Ryoo K, Kang KW, et al. Glutathione S-transferase Mu એપોપ્ટોસિસ સિગ્નલ-રેગ્યુલેટિંગ કિનેઝ 1. J Biol Chem ને દબાવીને સ્ટ્રેસ એક્ટિવેટેડ સિગ્નલોને મોડ્યુલેટ કરે છે. 2001;276:12749�12755.
88. Dorion S, Lambert H, Landry J. ગરમીના આંચકા દ્વારા p38 સિગ્નલિંગ પાથવેના સક્રિયકરણમાં Ask1 માંથી glutathione S-transferase Mu ના વિયોજનનો સમાવેશ થાય છે. જે બાયોલ કેમ. 2002;277:30792�30797.
89. એડલર V, Yin Z, Fuchs SY, Benezra M, Rosario L, et al. GSTp દ્વારા JNK સિગ્નલિંગનું નિયમન. EMBO જે. 1999;18:1321�1334.
90. Manevich Y, Feinstein SI, Fisher AB. એન્ટીઑકિસડન્ટ એન્ઝાઇમ 1-CYS પેરોક્સિરેડોક્સિનના સક્રિયકરણ માટે પીજીએસટી સાથે હેટરોડાઇમરાઇઝેશન દ્વારા મધ્યસ્થી ગ્લુટાથિઓનિલેશનની જરૂર છે. Proc Natl Acad Sci US A. 2004;101:3780�3785.
91. બંકર VW. મુક્ત રેડિકલ, એન્ટીઑકિસડન્ટો અને વૃદ્ધત્વ. મેડ લેબ સાય. 1992;49:299�312.
92. Mezzetti A, Lapenna D, Romano F, Costantini F, Pierdomenico SD, et al. પ્રણાલીગત ઓક્સિડેટીવ તણાવ અને વય અને માંદગી સાથે તેનો સંબંધ. J Am Geriatr Soc. 1996;44:823�827.
93. વ્હાઇટ ઇ, શેનોન જેએસ, પેટરસન આરઇ. વિટામિન અને વચ્ચેનો સંબંધ
કેલ્શિયમ પૂરક ઉપયોગ અને કોલોન કેન્સર. કેન્સર એપિડેમિઓલ બાયોમાર્કર્સ અગાઉના. 1997;6:769�774.
94. માસેલા આર, ડી બેનેડેટો આર, વેરી આર, ફાઇલસી સી, જીઓવાન્ની સી. જૈવિક પ્રણાલીઓમાં કુદરતી એન્ટીઑકિસડન્ટ સંયોજનોની નોવેલ મિકેનિઝમ્સ: ગ્લુટાથિઓન અને ગ્લુટાથિઓન-સંબંધિત ઉત્સેચકોની સંડોવણી. જે ન્યુટ્ર બાયોકેમ. 2005;16:577�586.
95. Curello S, Ceconi C, Bigoli C, Ferrari R, Albertini A, Guarnieri C. ઇસ્કેમિયા અને રિપરફ્યુઝન પછી કાર્ડિયાક ગ્લુટાથિઓન સ્થિતિમાં ફેરફાર. અનુભવ. 1985;41:42�43.
96. El-Agamey A, Lowe GM, McGarvey DJ, Mortensen A, Phillip DM, Truscott TG. કેરોટીનોઇડ રેડિકલ રસાયણશાસ્ત્ર અને એન્ટીઑકિસડન્ટ/પ્રો-ઑક્સિડન્ટ ગુણધર્મો. આર્ક બાયોકેમ બાયોફિઝ. 2004;430:37�48.
97. રાઇસ-ઇવાન્સ સીએ, સેમ્પસન જે, બ્રામલી પીએમ, હોલોવે ડીઇ. શા માટે આપણે વિવોમાં કેરોટીનોઇડ્સ એન્ટીઑકિસડન્ટ હોવાની અપેક્ષા રાખીએ છીએ? ફ્રી રેડિક રેસ. 1997;26:381�398.
98. નાઇલ્સ આરએમ. રેટિનોઇડ કીમોપ્રિવેન્શન અને કેન્સરની સારવારમાં સિગ્નલિંગ પાથવેઝ. Mutat Res. 2004;555:81�96.
99. ડોનાટો એલજે, નોય એન. રેટિનોઇક એસિડ દ્વારા સ્તન્ય પ્રાણીઓમાં ગર્ભમાં રહેલા બચ્ચાની રક્ષા માટેનું આચ્છાદન કાર્સિનોમા વૃદ્ધિનું દમન: પ્રોઆપોપ્ટોટિક જનીનો એ રેટિનોઇક એસિડ રીસેપ્ટર અને સેલ્યુલર રેટિનોઇક એસિડ-બંધનકર્તા પ્રોટીન II સિગ્નલિંગ માટે લક્ષ્ય છે. કેન્સર રે. 2005;65:8193�8199.
100. નિઝુમા એચ, નાકામુરા વાય, ઓઝાકી ટી, નાકાનિશી એચ, ઓહિરા એમ, એટ અલ. Bcl-2 એ ન્યુરોબ્લાસ્ટોમામાં રેટિનોઇક એસિડ-પ્રેરિત એપોપ્ટોટિક કોષ મૃત્યુ માટેનું મુખ્ય નિયમનકાર છે. ઓન્કોજીન. 2006;25:5046�5055.
101. ડાલ્ટન ટીપી, શર્ટઝર એચજી, પુગા એ. પ્રતિક્રિયાશીલ ઓક્સિજન દ્વારા જનીન અભિવ્યક્તિનું નિયમન. એન રેવ ફાર્માકોલ ટોક્સિકોલ. 1999;39:67�101.
102. સ્કેન્ડલિયોસ જેજી. ઓક્સિડેટીવ તણાવ માટે જીનોમિક પ્રતિભાવો. માં: મેયર્સ આરએ, એડ. મોલેક્યુલર સેલ બાયોલોજી અને મોલેક્યુલર મેડિસિનનો જ્ઞાનકોશ. વોલ્યુમ 5. 2જી આવૃત્તિ. Weinheim, જર્મની: Wiley-VCH; 2004: 489�512.
103. ઘોષ આર, મિશેલ ડીએલ. ટ્રાન્સક્રિપ્શન પરિબળ બંધનકર્તા પર પ્રમોટર તત્વોમાં ઓક્સિડેટીવ ડીએનએ નુકસાનની અસર. ન્યુક્લિક એસિડ્સ Res. 1999;27:3213�3218.
104. મેરીએટા સી, ગુલામ એચ, બ્રુક્સ પીજે. TATA બોક્સમાં એક જ 8, 50-cyclo-20-deoxyadenosine જખમ TATA બંધનકર્તા પ્રોટીનને બંધાતા અટકાવે છે અને વિવોમાં ટ્રાન્સક્રિપ્શનને મજબૂત રીતે ઘટાડે છે. DNA રિપેર (Amst). 2002;1:967�975.
105. જેક્સન એએલ, ચેન આર, લોએબ એલએ. માઇક્રોસેટેલાઇટ અસ્થિરતાનું ઇન્ડક્શન
ઓક્સિડેટીવ ડીએનએ નુકસાન દ્વારા. Proc Natl Acad Sci US A. 1998;95:12468�12473.
106. કેલ્ડેકોટ કેડબલ્યુ. સસ્તન પ્રાણી ડીએનએ સિંગલ-સ્ટ્રેન્ડ બ્રેક રિપેર દરમિયાન પ્રોટીન-પ્રોટીન ક્રિયાપ્રતિક્રિયાઓ. બાયોકેમ સોક ટ્રાન્સ. 2003;31:247�251.
107. કૂક એમએસ, ઇવાન્સ એમડી, ડીઝડારોગ્લુ એમ, લ્યુનેક જે. ઓક્સિડેટીવ ડીએનએ નુકસાન: મિકેનિઝમ્સ, મ્યુટેશન અને રોગ. FASEB જે. 2003; 17:1195�1214.
108. જોન્સ પીએલ, વોલ્ફ એપી. જનીન અભિવ્યક્તિ નક્કી કરવામાં ક્રોમેટિન સંસ્થા અને ડીએનએ મેથિલેશન વચ્ચેના સંબંધો. સેમિન કેન્સર બાયોલ. 1999;9:339�347.
109. ગીરોટ્ટી AW. લિપિડ પેરોક્સિડેશનની પદ્ધતિઓ. જે ફ્રી રેડિક બાયોલ મેડ. 1985;1:87�95.
110. સિયુ જીએમ, ડ્રેપર એચએચ. વિવો અને ઇન વિટ્રોમાં મેલોનાલ્ડીહાઇડનું ચયાપચય. લિપિડ્સ. 1982;17:349�355.
111. Esterbauer H, Koller E, Slee RG, Koster JF. ફ્લોરોસન્ટ ક્રોમોલિપિડ્સની રચનામાં લિપિડ-પેરોક્સિડેશન પ્રોડક્ટ 4-હાઈડ્રોક્સિનોનેનલની સંભવિત સંડોવણી. બાયોકેમ જે. 1986;239:405�409.
112. Hagihara M, Nishigaki I, Maseki M, Yagi K. માનવ સીરમના લિપોપ્રોટીન અપૂર્ણાંકમાં લિપિડ પેરોક્સાઇડ સ્તરોમાં વય-આધારિત ફેરફારો. જે ગેરોન્ટોલ. 1984;39:269�272.
113. કેલર જેએન, માર્ક આરજે, બ્રુસ એજે, બ્લેન્ક ઇ, રોથસ્ટીન જેડી, એટ અલ. 4- હાઇડ્રોક્સિનોનેનલ, મેમ્બ્રેન લિપિડ પેરોક્સિડેશનનું એલ્ડીહાઇડિક ઉત્પાદન, સિનેપ્ટોસોમ્સમાં ગ્લુટામેટ પરિવહન અને મિટોકોન્ડ્રીયલ કાર્યને નબળી પાડે છે. ન્યુરોસાયન્સ. 1997;806:85�96.
114. Uchida K, Shiraishi M, Naito Y, Torii Y, Nakamura Y, Osawa T. લિપિડ પેરોક્સિડેશનના અંતિમ ઉત્પાદન દ્વારા સ્ટ્રેસ સિગ્નલિંગ પાથવેઝનું સક્રિયકરણ. 4-hydroxy-2-nonenal એ અંતઃકોશિક પેરોક્સાઇડ ઉત્પાદનનું સંભવિત પ્રેરક છે. જે બાયોલ કેમ. 1999;274:2234�2242.
115. Suc I, Meilhac O, Lajoie-Mazenc I, Vandaele J, Jurgens G, Salvayre R, Negre-Salvayre A. ઓક્સિડાઇઝ્ડ LDL દ્વારા EGF રીસેપ્ટરનું સક્રિયકરણ. FASEB જે. 1998; 12:665�671.
116. ત્સુકાગોશી એચ, કવાટા ટી, શિમિઝુ વાય, ઇશિઝુકા ટી, ડોબાશી કે, મોરી એમ. 4-હાઇડ્રોક્સી-2-નોનેનલ એપિડર્મલ વૃદ્ધિ પરિબળ રીસેપ્ટર-લિંક્ડ એક્સ્ટ્રા સેલ્યુલર સિગ્નલના સક્રિયકરણ દ્વારા IMR-90 માનવ ફેફસાના ફાઇબ્રોબ્લાસ્ટ દ્વારા ફાઇબ્રોનેક્ટીન ઉત્પાદનમાં વધારો કરે છે. રેગ્યુલેટેડ કિનાઝ p44/42 પાથવે. ટોક્સિકોલ એપ્લ ફાર્માકોલ. 2002;184:127�135.
117. મોન્ટુચી પી, કોલિન્સ જેવી, સિયાબેટોની જી, લેઝેરી એન, કોરાડી એમ, ખારીટોનોવ એસએ, બાર્નેસ પીજે. સીઓપીડી અને તંદુરસ્ત ધુમ્રપાન કરનારા દર્દીઓમાં ફેફસાના ઓક્સિડેટીવ તાણના ઇન વિવો બાયોમાર્કર તરીકે 8-આઇસોપ્રોસ્ટેન શ્વાસ બહાર કાઢે છે. એમ જે રેસ્પિર ક્રિટ કેર મેડ. 2000;162:1175�1177.
118. મોરિસન ડી, રહેમાન I, લેનાન એસ, મેકની ડબલ્યુ. ઉપકલા અભેદ્યતા, બળતરા, અને ધુમ્રપાન કરનારાઓની હવાની જગ્યામાં ઓક્સિડન્ટ તણાવ. એમ જે રેસ્પિર ક્રિટ કેર મેડ. 1999;159:473�479.
119. Nowak D, Kasielski M, Antczak A, Pietras T, Bialasiewicz P. સ્થિર ક્રોનિક અવરોધક પલ્મોનરી રોગ ધરાવતા દર્દીઓના નિવૃત્ત શ્વાસ કન્ડેન્સેટમાં થિયોબાર્બિટ્યુરિક એસિડ-પ્રતિક્રિયાશીલ પદાર્થો અને હાઇડ્રોજન પેરોક્સાઇડની સામગ્રીમાં વધારો: કિંગમોસિગરની કોઈ નોંધપાત્ર અસર નથી. રેસ્પિર મેડ. 1999;93:389�396.
120. કેલી એફજે, મુડવે IS. હવા-ફેફસાના ઇન્ટરફેસ પર પ્રોટીન ઓક્સિડેશન. એમિનો એસિડ. 2003;25:375�396.
121. ડીન આરટી, રોબર્ટ્સ સીઆર, જેસપ ડબલ્યુ. મુક્ત રેડિકલ દ્વારા એક્સ્ટ્રા સેલ્યુલર અને ઇન્ટ્રાસેલ્યુલર પોલિપેપ્ટાઇડ્સનું ફ્રેગમેન્ટેશન. પ્રોગ ક્લિન બાયોલ રેસ. 1985;180:341�350.
122. કેક આરજી. પ્રોટીનમાં મેથિઓનાઇન ઓક્સિડેશન માટે ચકાસણી તરીકે ટી-બ્યુટીલ હાઇડ્રોપેરોક્સાઇડનો ઉપયોગ. ગુદા બાયોકેમ. 1996;236:56�62.
123. ડેવિસ કેજે. ઓક્સિજન રેડિકલ દ્વારા પ્રોટીનને નુકસાન અને અધોગતિ. I. સામાન્ય પાસાઓ. જે બાયોલ કેમ. 1987;262:9895�9901.
124. સ્ટેડટમેન ER. પ્રોટીનનું મેટલ આયન-ઉત્પ્રેરિત ઓક્સિડેશન: બાયોકેમિકલ મિકેનિઝમ અને જૈવિક પરિણામો. ફ્રી રેડિક બાયોલ મેડ.
1990;9:315�325.
125. Fucci L, Oliver CN, Coon MJ, Stadtman ER. મિશ્ર-કાર્ય ઓક્સિડેશન પ્રતિક્રિયાઓ દ્વારા મુખ્ય મેટાબોલિક ઉત્સેચકોનું નિષ્ક્રિયકરણ: પ્રોટીન ટર્નઓવર અને વૃદ્ધત્વમાં સંભવિત સૂચિતાર્થ. Proc Natl Acad Sci US A. 1983;80:1521�1525.
126. Stadtman ER, Moskovitz J, Levine RL. પ્રોટીનના મેથિઓનાઇન અવશેષોનું ઓક્સિડેશન: જૈવિક પરિણામો. એન્ટિઓક્સિડ રેડોક્સ સિગ્નલ. 2003;5:577�582.
127. સ્ટેડટમેન ER, લેવિન આરએલ. પ્રોટીનમાં મુક્ત એમિનો એસિડ અને એમિનો એસિડ અવશેષોનું મુક્ત રેડિકલ-મધ્યસ્થ ઓક્સિડેશન. એમિનો એસિડ. 2003;25:207�218.
128. સ્ટેડટમેન ER. વૃદ્ધત્વ અને વય-સંબંધિત રોગોમાં પ્રોટીન ઓક્સિડેશન. એન NY Acad Sci. 2001;928:22�38.
129. શેક્ટર E. જૈવિક નમૂનાઓમાં પ્રોટીન ઓક્સિડેશનનું પ્રમાણીકરણ અને મહત્વ. ડ્રગ મેટાબ રેવ. 2000;32:307�326.
130. Poli G, Leonarduzzi G, Biasi F, Chiarpotto E. ઓક્સિડેટીવ સ્ટ્રેસ અને સેલ સિગ્નલિંગ. કર મેડ કેમ. 2004;11:1163�1182.
131. ન્યુફેલ્ડ જી, કોહેન ટી, ગેંગરીનોવિચ એસ, પોલ્ટોરક ઝેડ. વેસ્ક્યુલર એન્ડોથેલિયલ ગ્રોથ ફેક્ટર (VEGF) અને તેના રીસેપ્ટર્સ. FASEB જે. 1999; 13:9�22.
132. સુંદરેસન એમ, યુ ઝેડએક્સ, ફેરન્સ વીજે, સલ્સિનર ડીજે, ગુટકાઇન્ડ જેએસ, એટ અલ. Rac1 દ્વારા ફાઇબ્રોબ્લાસ્ટ્સમાં પ્રતિક્રિયાશીલ-ઓક્સિજન પ્રજાતિઓનું નિયમન. બાયોકેમ જે. 1996;318:379�382.
133. સન ટી, ઓબરલી એલડબ્લ્યુ. ટ્રાન્સક્રિપ્શનલ એક્ટિવેટર્સનું રેડોક્સ નિયમન. ફ્રી રેડિક બાયોલ મેડ. 1996;21:335�348.
134. Klatt P, Molina EP, De Lacoba MG, Padilla CA, Martinez-Galesteo E, Barcena JA, Lamas S. Redox રેગ્યુલેશન ઓફ c-Jun DNA બંધનકર્તા S-glutathiolation દ્વારા. FASEB જે. 1999; 13:1481�1490.
135. Reynaert NL, Ckless K, Guala AS, Wouters EF, van der Vliet A, Janssen Heininger
YM. ગ્લુટારેડોક્સિન-1 ઉત્પ્રેરિત સિસ્ટીન ડેરિવેટાઇઝેશન પછી એસ-ગ્લુટાથિઓનિલેટેડ પ્રોટીનની પરિસ્થિતિમાં શોધ. બાયોચિમ બાયોફિઝ એક્ટા. 2006;1760:380�387.
136. Reynaert NL, Wouters EF, Janssen-Heininger YM. ગ્લુટેરેડોક્સિન-1નું મોડ્યુલેશન
એલર્જિક એરવે રોગના માઉસ મોડેલમાં અભિવ્યક્તિ. એમ જે રેસ્પિર સેલ મોલ બાયોલ. 2007;36:147�151.
137. Filomeni G, Rotilio G, Ciriolo MR. સેલ સિગ્નલિંગ અને ગ્લુટાથિઓન રેડોક્સ સિસ્ટમ. બાયોકેમ ફાર્માકોલ. 2002;64:1057�1064.
138. પાંડે વી, રામોસ એમજે. પરમાણુ પરિબળ-કપ્પા બી અને અન્ય સેલ્યુલર પ્રોટીન દ્વારા 15-ડીઓક્સીડેલ્ટા (12,14) પ્રોસ્ટાગ્લાન્ડિન J(2) ની પરમાણુ ઓળખ. Bioorg Med Chem Lett. 2005;15:4057�4063.
139. પર્કિન્સ એનડી. NF-kappaB અને IKK ફંક્શન સાથે સેલ-સિગ્નલિંગ પાથવેને એકીકૃત કરવું. નેટ રેવ મોલ સેલ બાયોલ. 2007;8:49�62.
140. ગિલમોર ટીડી. NF-kappaB નો પરિચય: ખેલાડીઓ, માર્ગો, દ્રષ્ટિકોણ. ઓન્કોજીન. 2006;25:6680�6684.
141. હિરોટા કે, મુરાતા એમ, સચી વાય, નાકામુરા એચ, ટેકયુચી જે, મોરી કે, યોડોઈ જે. સાયટોપ્લાઝમ અને ન્યુક્લિયસમાં થિયોરેડોક્સિનની વિશિષ્ટ ભૂમિકાઓ. ટ્રાન્સક્રિપ્શન ફેક્ટર NF-kappaB ના રેડોક્સ નિયમનની બે-પગલાની પદ્ધતિ. જે બાયોલ કેમ. 1999;274:27891�27897.
142. વોર્ડ PA. તીવ્ર ફેફસાની ઇજામાં પૂરક, કેમોકાઇન્સ અને નિયમનકારી સાઇટોકીન્સની ભૂમિકા. એન NY Acad Sci. 1996;796:104�112.
143. અકીરા એસ, કિશિમોટો એ. NF-IL6 અને NF-kB સાયટોકાઇન જનીન નિયમનમાં. એડ્વ ઇમ્યુનોલ. 1997;65:1�46.
144. મેયર એમ, શ્રેક આર, બેઉર્લે પીએ. H2O2 અને એન્ટીઑકિસડન્ટો અખંડ કોષોમાં NF-kappa B અને AP-1 ના સક્રિયકરણ પર વિપરીત અસરો ધરાવે છે: AP-1 ગૌણ એન્ટીઑકિસડન્ટ-પ્રતિભાવ પરિબળ તરીકે. EMBO જે. 1993;12:2005�2015.
145. એબેટ સી, પટેલ એલ, રાઉશર એફજે, કુરન ટી. રેડોક્સ રેગ્યુલેશન ઓફ ફોસ અને જૂન ડીએનએ-બાઈન્ડિંગ એક્ટિવિટી ઇન વિટ્રો. વિજ્ઞાન. 1990;249:1157�1161.
146. ગેલ્ટર ડી, મિહમ એસ, ડ્રોજ ડબલ્યુ. ન્યુક્લિયર ટ્રાન્સક્રિપ્શન ફેક્ટર્સ kB અને એક્ટિવેટર પ્રોટીન-1 પર ગ્લુટાથિઓન ડિસલ્ફાઇડની વિશિષ્ટ અસરો. યુર જે બાયોકેમ. 1994;221:639�648.
147. Hirota K, Matsui M, Iwata S, Nishiyama A, Mori K, Yodoi J. AP-1 ટ્રાન્સક્રિપ્શનલ પ્રવૃત્તિ થિયોરેડોક્સિન અને રેફ-1 વચ્ચેના સીધા જોડાણ દ્વારા નિયંત્રિત થાય છે. Proc Natl Acad Sci US A. 1997;94: 3633�3638.
IFMનું ફાઇન્ડ અ પ્રેક્ટિશનર ટૂલ એ ફંક્શનલ મેડિસિનનું સૌથી મોટું રેફરલ નેટવર્ક છે, જે દર્દીઓને વિશ્વમાં ક્યાંય પણ ફંક્શનલ મેડિસિન પ્રેક્ટિશનર્સ શોધવામાં મદદ કરવા માટે બનાવવામાં આવ્યું છે. IFM સર્ટિફાઇડ પ્રેક્ટિશનર્સ શોધ પરિણામોમાં પ્રથમ સૂચિબદ્ધ થાય છે, તેઓ કાર્યકારી દવામાં વ્યાપક શિક્ષણ આપે છે.